

Resumen
El envejecimiento provoca una pérdida progresiva de la función del sistema inmune que nos vuelve más vulnerables al cáncer y a las infecciones, disminuye nuestra respuesta a las vacunas, y aumenta el riesgo de padecer enfermedades autoinmunes. Evidencias recientes indican que las alteraciones en los linfocitos T que se acumulan con la edad provocan una inflamación crónica que precipita patologías propias del envejecimiento. En nuestro laboratorio hemos generado unos ratones que a edad temprana presentan un sistema inmune envejecido. Estos ratones tienen la esperanza de vida reducida a la mitad, presentan senescencia prematura en diferentes órganos y patología cardiovascular, metabólica y sarcopenia. Así, hemos demostrado que el envejecimiento de las células T acelera la multimorbilidad asociada a la edad. Entender cómo el sistema inmune contribuye a esta multimorbilidad asociada a la edad es un desafío urgente y prioritario que nos permitirá desarrollar estrategias terapéuticas para tener un envejecimiento saludable. En este sentido, la inmunoterapia podría representar una nueva estrategia no solo para combatir el cáncer sino para retrasar las enfermedades relacionadas con el envejecimiento.
Abstract
Aging of the immune system is characterized by a progressive loss of function, which limits vaccine effectiveness and is responsible for the increase in the susceptibility to infections, cancer and autoimmune diseases observed in elderly population. Recent evidences suggest that intrinsic alterations accumulated in T lymphocytes with age actively contribute to the state of chronic inflammation which underlies age-associated pathologies. We have accelerated the aging process in mice by inducing a mitochondrial dysfunction exclusively in T cells. These mice showed signs of systemic premature senescence and multimorbidity, evidencing that alterations in T cell functionality are sufficient to accelerate a whole-organism aging phenotype. Thus, T cell aging plays a crucial role in systemic deterioration and may be involved in neurodegenerative, cardiovascular and metabolic diseases whose incidence drastically increases with age. Understanding how the immune system contributes to age-associated multimorbidity is an urgent challenge, which could lead to the development of new therapeutic strategies destined to achieve a healthier aging. In this regard, immunotherapy is emerging as a new and promising technique aimed to delay age-associated diseases.
Palabras clave: Envejecimiento; Linfocitos T; Senescencia; Inmunoterapia; Mitocondria.
Keywords: Aging; T cells; Senescence; Immunotherapy; Mitochondria.
INTRODUCCIÓN
Con la edad las células acumulan daños en el ADN y en sus orgánulos que comprometen su función y las llevan a un estado de senescencia. Las células senescentes se van acumulando progresivamente en los tejidos y contribuyen activamente al deterioro tisular y al envejecimiento (1). Las células senescentes se caracterizan por presentar daño en el ADN, disfunción mitocondrial y una pérdida de su capacidad de división, que lleva a un estado de arresto del ciclo celular. Además, adquieren un fenotipo denominado SASP (Senescence-associated Secretory Phenotype) caracterizado por la secreción de un amplio repertorio de citoquinas proinflamatorias (e.g. IL6, CXCL1, IL8, IL1β), que comprometen la funcionalidad del tejido en el que se encuentran. Este fenotipo SASP fomenta una inflamación crónica de bajo grado que está asociada a la aparición de diferentes enfermedades propias del envejecimiento en un proceso conocido como “inflammaging” (2).
Los linfocitos T, como cualquier otra célula del organismo, también se ven afectados con la edad, recapitulando ciertas características propias de células senescentes. Se estima que existen cientos de millones linfocitos T circulando por nuestro cuerpo, inspeccionando cada uno de nuestros órganos como centinelas silenciosos (4). Cuando reconocen un antígeno extraño mediante su TCR, los linfocitos T se activan, se dividen rápidamente y se diferencian en distintas subpoblaciones de linfocitos T CD4+ efectores (Th1, Th2, Th17 o Treg) o citotóxicos (CD8+) para coordinar una respuesta inmune y eliminar aquellas células que suponen una amenaza para la homeostasis del organismo, como es el caso de células infectadas, tumorales o senescentes. Sin embargo, el envejecimiento y la senescencia de estas células del sistema inmune afecta negativamente a su función.
Los linfocitos T se generan y maduran en el timo, un órgano linfoide localizado entre el corazón y el esternón. Allí, mediante procesos de selección positiva y negativa, son entrenados para distinguir los antígenos propios de los ajenos, asegurando así un correcto funcionamiento del sistema inmune. El timo es el primer órgano que se deteriora con la edad, teniendo una actividad máxima desde que nacemos hasta la adolescencia. A partir de esa edad comienza a decaer, de manera que a los 20 años apenas hay actividad tímica y a los cuarenta años prácticamente todo el timo ha sido reemplazado por tejido adiposo (Figura 1). Como consecuencia de esta atrofia tímica, dejan de generarse nuevos linfocitos T. Por ello, el mantenimiento de los niveles adecuados de estas células depende, exclusivamente, de células preexistentes que sufren una gran presión replicativa, pues con cada división celular se acumulan daños citogenéticos que comprometen su funcionalidad, pudiendo perder la capacidad de distinguir lo propio de lo ajeno. Gracias a la técnica de secuenciación del ARN mensajero de células individuales hemos aprendido que durante el envejecimiento desaparecen las células T naïve y se van acumulando células T memoria, muchas de ellas con características de células senescentes o exhaustas (Figura 1). Al contrario que las células T senescentes que se caracterizan por el fenotipo SASP, las células T exhaustas dejan de producir citoquinas y son incapaces de responder a estímulos inflamatorios (3). Todos estos cambios en el repertorio de células T aumentan la vulnerabilidad a las infecciones, así como la predisposición a padecer cáncer y enfermedades autoinmunes, reducen la respuesta a la vacunación y favorecen la aparición del inflammaging (Figura 1) (6).
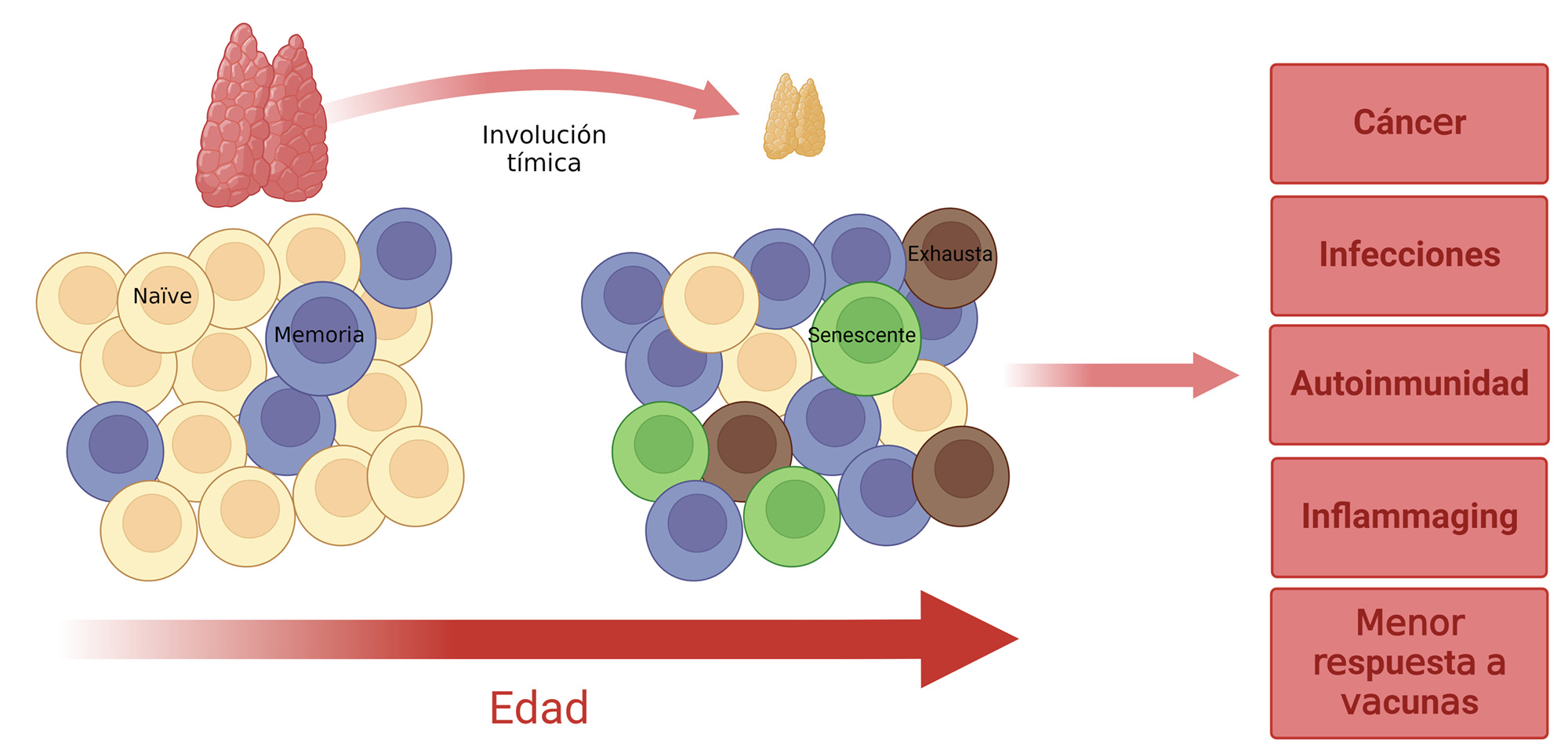
DESINCRONIZANDO LA EDAD DEL SISTEMA INMUNE DE LA DEL RESTO DE TEJIDOS
La hipótesis de nuestro laboratorio es que el declive de la función del sistema inmune con la edad no solo nos hace mucho más vulnerables a las infecciones y al cáncer, sino que, además, aumenta el riesgo de padecer patologías propias del envejecimiento como enfermedades cardiovasculares, neurodegenerativas y metabólicas. Para demostrar nuestra hipótesis, hemos generado un modelo de ratón cuyos linfocitos T, CD4+ y CD8+, envejecen de forma prematura debido a una disfunción mitocondrial.
Las mitocondrias son los orgánulos responsables de generar energía en forma de ATP mediante el proceso bioquímico de fosforilación oxidativa que recae en la correcta función de 4 complejos multiproteicos (complejos I al IV) que conforman la cadena de transporte de electrones y generan un gradiente electroquímico empleado por la ATP sintasa (complejo V) para la síntesis de ATP. Las proteínas que forman parte de estos complejos están codificadas tanto en el ADN nuclear como en el ADN mitocondrial (ADNmt) (7).
A diferencia del ADN nuclear, que cuenta con múltiples mecanismos de reparación y corrección de errores durante su replicación, el ADNmt apenas dispone de sistemas de protección debido a su origen procariota y está altamente expuesto a estrés oxidativo. Por ello, con la edad, el ADNmt va acumulando daño, siendo la disfunción mitocondrial una de las primeras marcas del envejecimiento celular (1).
Para investigar las consecuencias del deterioro del sistema inmune con la edad, hemos adelantado la disfunción mitocondrial que ocurre normalmente en todos los tejidos durante el envejecimiento, exclusivamente en los linfocitos T. Para ello, hemos generado un modelo de ratón con una depleción del gen que codifica para el factor de transcripción mitocondrial A (Tfam) exclusivamente en los linfocitos T. TFAM es una de las proteínas más importantes para la estabilidad, replicación y transcripción del ADNmt por lo que su depleción reduce el contenido de ADNmt, y de las 13 subunidades de la cadena de transporte de electrones allí codificadas generando un mal funcionamiento de la mitocondria (Figura 2). La depleción de TFAM en el modelo fue inducida empleando el sistema de recombinación Cre-loxP, a partir del cruce de ratones Tfamfl/fl generados en el laboratorio del Prof. N. G. Larsson, con ratones que poseen la recombinasa Cre bajo el promotor de CD4 (CD4Cre). Mediante esta estrategia, se consigue la deleción del gen en las primeras etapas de desarrollo de las células T, de manera que la expresión de Tfam está impedida tanto en linfocitos T CD4+ como CD8+. Los ratones Tfamfl/fl CD4Cre se desarrollan con normalidad y tienen un número similar de linfocitos CD4+ y CD8+ en el bazo y en los ganglios linfáticos en comparación con los ratones control, lo que indica que Tfam no se requiere durante el desarrollo de las células T. Así los linfocitos T con una depleción de Tfam presentan una severa disfunción mitocondrial (8).
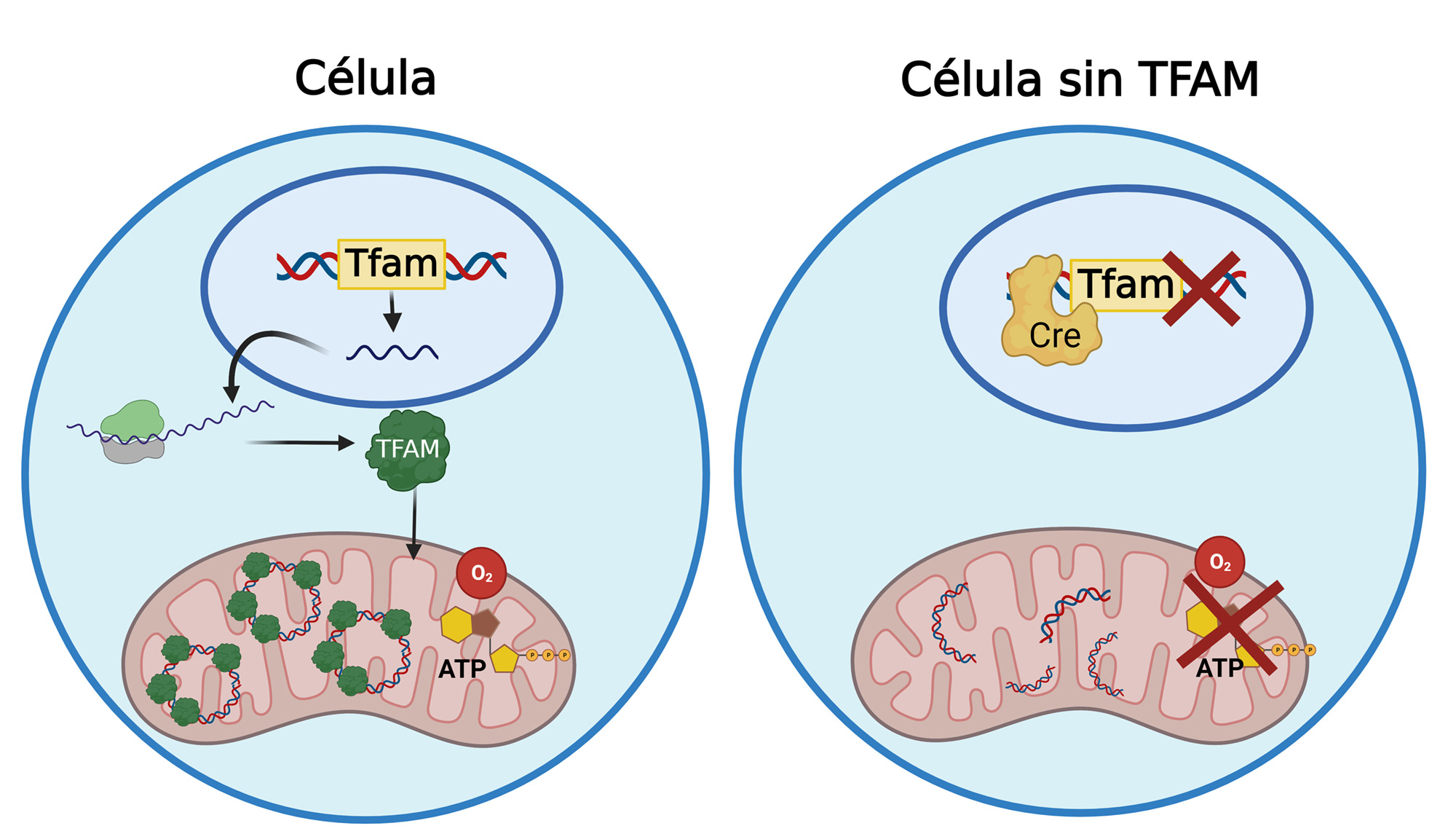
Esta alteración de la función mitocondrial en los linfocitos T es suficiente para recapitular varias de las señales de envejecimiento del sistema inmune: disminución de la proporción de linfocitos T naïve y aumento de linfocitos T de memoria que presentan fenotipos pro-inflamatorios y senescentes, así como mayor vulnerabilidad frente a cáncer o infecciones. De hecho, al infectar ratones con el virus Ectromelia, virus de la viruela de ratón, el 100% de los ratones jóvenes sobrevivieron mientras que el 100% de los ratones viejos y de los ratones jóvenes Tfamfl/fl CD4Cre sucumbieron a la infección. Estos resultados demuestran que la disfunción mitocondrial inducida en los linfocitos T produce un envejecimiento prematuro del sistema inmune tanto a nivel molecular como funcional.
LOS LINFOCITOS T ENVEJECIDOS ACELERAN LA SENESCENCIA DE LOS TEJIDOS Y LA MULTIMORBILIDAD ASOCIADA A LA EDAD
Además de la alteración en la función del sistema inmune, los ratones Tfamfl/fl CD4Cre presentaron una reducción a la mitad de su esperanza de vida, acompañada de la aparición acelerada de marcas de senescencia en otros tejidos como riñón, hígado y tejido adiposo, y una multimorbilidad prematura, es decir, a edad temprana estos ratones presentaban ya enfermedades propias del envejecimiento, tales como alteraciones cardiovasculares, sarcopenia y deterioro cognitivo (9). En concordancia con estos resultados, un laboratorio independiente demostró que el trasplante de un sistema inmune envejecido de forma prematura mediante daño genotóxico en el ADN nuclear también es capaz de inducir senescencia en el resto de los tejidos (10). Estos datos sugieren que el envejecimiento del sistema inmune tiene consecuencias a nivel sistémico, apoyando la hipótesis de nuestro laboratorio de que el sistema inmune envejecido favorece la acumulación de células senescentes en otros tejidos, acelerando el envejecimiento del resto del organismo.
Los mecanismos moleculares por los cuales las células T pueden contribuir al envejecimiento han sido y son objeto de estudio por nuestro laboratorio y por otros laboratorios. Se han postulado distintos mecanismos que explican este proceso. Por un lado, los linfocitos T envejecidos adquieren un fenotipo proinflamatorio senescente de tipo Th1, que se caracteriza por la secreción de citoquinas como TNF e IFNγ (8). Estas citoquinas son capaces de inducir senescencia cuando se cultivan in vitro (11), sugiriendo que los linfocitos Th1 pueden inducir senescencia en distintos tejidos de forma paracrina (Figura 3). Alternativamente, los linfocitos T envejecidos pueden perder su capacidad protectora para detectar y eliminar células senescentes (5), favoreciendo la acumulación de células senescentes de forma indirecta. Otra posibilidad es que los linfocitos T deteriorados pierdan el control sobre la microbiota, favoreciendo la aparición de una disbiosis intestinal que puede desencadenar permeabilidad intestinal. Esto favorecería la translocación de bacterias a la sangre generando una inflamación crónica subclínica que deteriora el funcionamiento de los tejidos y favorece la acumulación de células senescentes (12).
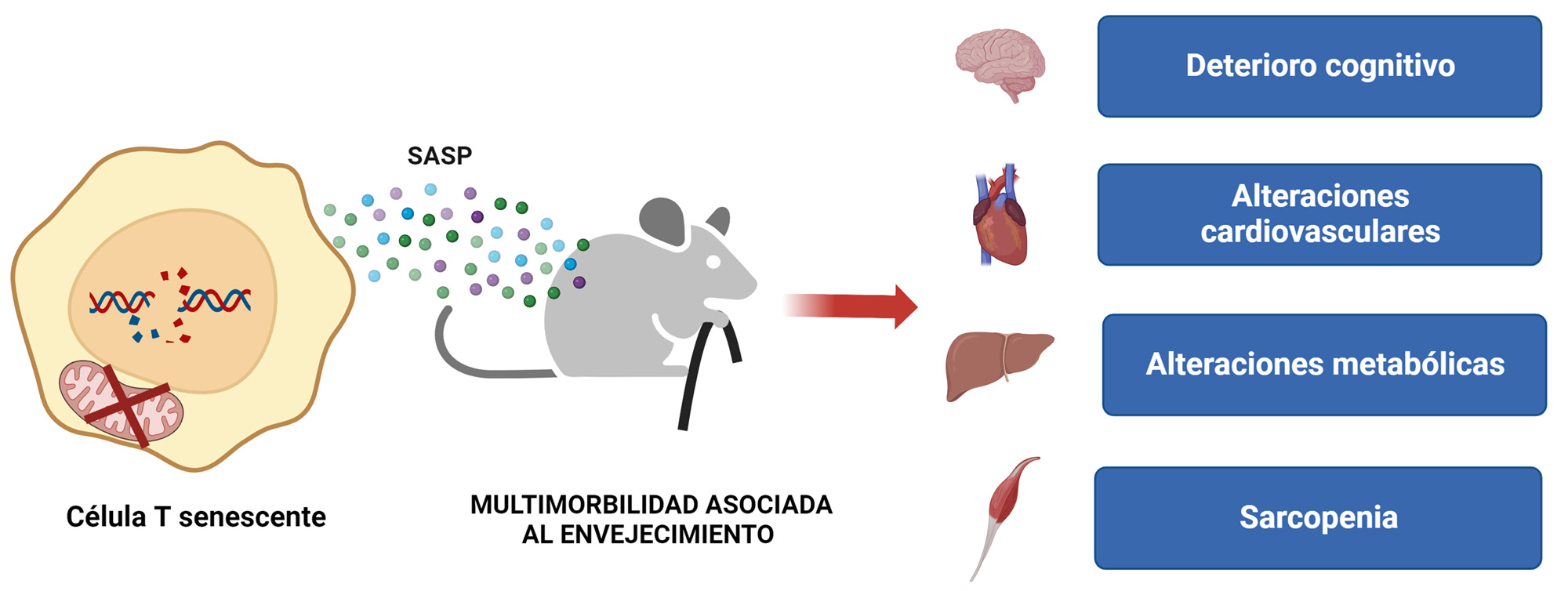
En resumen, estos resultados sitúan la función del sistema inmune, y en concreto a los linfocitos T, en el centro de nuestra salud. No solo son esenciales para prevenir enfermedades infecciosas, autoinmunes o el cáncer, sino que además tienen un papel clave en el control de la calidad y la velocidad a la que envejecemos. Todo ello invita a plantear la posibilidad de rejuvenecer el sistema inmune, o prevenir su deterioro, y con ello aminorar o retrasar alguna de las manifestaciones del envejecimiento.
INMUNOTERAPIA: LA NUEVA HERRAMIENTA TERAPÉUTICA PARA LAS ENFERMEDADES RELACIONADAS CON LA EDAD
En los últimos años, la inmunología ha revolucionado el campo de la oncología gracias a la inmunoterapia y al uso de los inhibidores de la regulación negativa del sistema inmune (anti-PD-1) y las células T con receptor de antígeno quimérico (CAR-T).
Los Drs. James Allison y Tasuku Honjo han sido reconocidos por la comunidad científica con el Premio Nobel de Fisiología o Medicina de 2018 por su descubrimiento de terapias del cáncer para inhibir la regulación negativa del sistema inmune. Gracias a sus investigaciones, hemos aprendido cómo eliminar los frenos de un sistema inmune agotado mediante el uso de reguladores de puntos de control inmunitarios para reactivar las células T exhaustas y generar una respuesta inmune más efectiva contra el cáncer. Esta estrategia ha supuesto un gran avance en el tratamiento del cáncer y también podría tener cabida en el campo de la gerociencia y la longevidad. De hecho, evidencias en modelos murinos experimentales de Alzheimer, sugieren que el uso de anticuerpos anti-PD-1 puede retrasar esta patología al activar el sistema inmune (13,14). Sin embargo, el bloqueo de PD-1 aumenta el riesgo de accidente cardiovascular al promover la ateroesclerosis, ya que la vía de señalización de PD-1 está implicada en el bloqueo de rutas proinflamatorias y proaterogénicas en células T (15). Todo ello nos lleva a pensar que otra posible estrategia frente al envejecimiento, y a determinadas enfermedades asociadas a él, podría ser emplear una aproximación opuesta a la utilizada en el tratamiento del cáncer, es decir, en vez de reactivar a las células T exhaustas, trataría de agotar a las células T senescentes.
Otra aproximación terapéutica propone diseñar estrategias que eliminen a las células T patogénicas, inflamatorias o senescentes. Se ha observado que la vacunación con el antígeno CD153 en ratón, reduce la acumulación de células T senescentes en el tejido adiposo y mejora los parámetros metabólicos (16). Alternativamente, desde el laboratorio del Dr. Lowe han diseñado unas células T modificadas genéticamente que expresan un receptor de antígeno quimérico (células CART), que les permite reconocer y eliminar específicamente las células senescentes que se acumulan en distintos tejidos. Estas células están dirigidas contra un marcador de superficie que se expresa principalmente en células senescentes, el receptor activador del plasminógeno de tipo uroquinasa (uPAR). La transferencia de las células CART específicas eliminó eficazmente las células senescentes en ratones con fibrosis hepática y aumentó la supervivencia en ratones con adenocarcinoma de pulmón (17). En la misma línea, se ha conseguido reducir la fibrosis cardiaca mediante la transferencia de células CART contra la proteína de activación de fibroblastos (FAP) (18).
A pesar de su alta eficacia para la eliminación de células senescentes demostrada en modelos experimentales, la producción de células CART es costosa y su durabilidad es baja una vez inyectadas en el paciente, por lo que han surgido estrategias todavía más sofisticadas destinadas a modificar los linfocitos T directamente in vivo. Para ello, se han diseñado nanopartículas lipídicas para transportar el ARN mensajero codificante del receptor de antígeno quimérico (CAR) dirigido a la FAP. Estas nanopartículas dirigidas específicamente a los linfocitos T fueron inyectadas en ratones, dando como resultado una expresión transitoria de CAR-anti-FAP en las células T, dotándolas de la capacidad de eliminar los fibroblastos activados y aliviar la fibrosis cardíaca en un modelo de ratón de fallo cardíaco (19).
CONCLUSIÓN
Debido al incremento de la esperanza de vida y la caída de la natalidad, nuestra sociedad está cada vez más envejecida. Se calcula que en 2050 más del 40% de la población tendrá más de 60 años. A partir de esa edad, el riesgo de multimorbilidad se incrementa exponencialmente. Por ello, entender por qué la edad nos vuelve más vulnerables a todas estas patologías es clave para el desarrollo de terapias que puedan retrasar la aparición de enfermedades propias del envejecimiento. En este sentido, el papel central del sistema inmune, y en concreto de los linfocitos T, en el desarrollo de la multimorbilidad asociada a la edad, hace que conocer los mecanismos moleculares detrás de dicho proceso sea desafío urgente y prioritario que nos permitirá desarrollar estrategias terapéuticas para tener un envejecimiento más saludable.
El potencial terapéutico de la inmunoterapia va más allá del tratamiento del cáncer. Su relevancia para el tratamiento de patologías relacionadas con la edad, como la fibrosis o la pérdida cognitiva, está empezando a despertar interés en la comunidad científica. Es posible que, durante la próxima década, la inmunoterapia comience a utilizarse para tratar los trastornos relacionados con el envejecimiento.
AGRADECIMIENTOS
Este trabajo ha sido financiado por el Fondo de Investigación Sanitaria del Instituto de Salud Carlos III (PI19/855), el Fondo para el Desarrollo Regional Europeo (ERDF) y la Comisión Europea por H2020-EU.1.1, el proyecto del Consejo de Investigación Europeo ERC-2016-StG 715322-EndoMitTalk y el proyecto sinérgico de la Comunidad de Madrid Y2020/Bio-6350 NutriSION-CM. El trabajo de M.M. está financiado por el Consejo Superior de Investigaciones Científicas (CSIC). El trabajo de S.D.P. está financiado por un contrato dentro del proyecto sinérgico de la Comunidad de Madrid Y2020/Bio-6350 NutriSION-CM. El trabajo de J.I.E.L. está financiado por la beca de Formación de Profesorado Universitario FPU20/0466 del Ministerio de Ciencia, Innovación y Universidades (España). Las figuras de este artículo han sido desarrolladas en biorender.com.
BIBLIOGRAFÍA
- López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153(6):1194-1217.
- Franceschi C, Garagnani P, Vitale G, Capri M, Salvioli S. Inflammaging and ‘Garb-aging’. Trends Endocrinol Metabolism. 2017;28(3):199-212.
- Elyahu Y, Hekselman I, Eizenberg-Magar I, et al. Aging promotes reorganization of the CD4 T cell landscape toward extreme regulatory and effector phenotypes. Sci Adv. 2019;5(8):eaaw8330.
- Jenkins MR, La Gruta NL, Doherty PC, Trapani JA, Turner SJ, Waterhouse NJ. Visualizing CTL activity for different CD8+ effector T cells supports the idea that lower TCR/epitope avidity may be advantageous for target cell killing. Cell Death Differ. 2009;16(4):537-542.
- Ovadya Y, Landsberger T, Leins H, et al. Impaired immune surveillance accelerates accumulation of senescent cells and aging. Nat Commun. 2018;9(1):5435.
- Mittelbrunn M, Kroemer G. Hallmarks of T cell aging. Nat Immunol. 2021;22(6):687-698.
- Friedman JR, Nunnari J. Mitochondrial form and function. Nature. 2014;505(7483):335-343.
- Baixauli F, Acín-Pérez R, Villarroya-Beltrí C, et al. Mitochondrial Respiration Controls Lysosomal Function during Inflammatory T Cell Responses. Cell Metab. 2015;22(3):485-498.
- Desdín-Micó G, Soto-Heredero G, Aranda JF, et al. T cells with dysfunctional mitochondria induce multimorbidity and premature senescence. Science. 2020;368(6497):1371-1376.
- Yousefzadeh MJ, Flores RR, Zhu Y, et al. An aged immune system drives senescence and ageing of solid organs. Nature. 2021;594(7861):100-105.
- Braumüller H, Wieder T, Brenner E, et al. T-helper-1-cell cytokines drive cancer into senescence. Nature. 2013;494(7437):361-365.
- Carrasco E, Gómez de las Heras MM, Gabandé-Rodríguez E, Desdín-Micó G, Aranda JF, Mittelbrunn M. The role of T cells in age-related diseases. Nat Rev Immunol. 2021;1-15.
- Baruch K, Deczkowska A, Rosenzweig N, et al. PD-1 immune checkpoint blockade reduces pathology and improves memory in mouse models of Alzheimer’s disease. Nat Med. 2016;22(2):135-137.
- Rosenzweig N, Dvir-Szternfeld R, Tsitsou-Kampeli A, et al. PD-1/PD-L1 checkpoint blockade harnesses monocyte-derived macrophages to combat cognitive impairment in a tauopathy mouse model. Nat Commun. 2019;10(1):465.
- Drobni ZD, Alvi RM, Taron J, et al. Association Between Immune Checkpoint Inhibitors With Cardiovascular Events and Atherosclerotic Plaque. Circulation. 2020;142(24):2299-2311.
- Yoshida S, Nakagami H, Hayashi H, et al. The CD153 vaccine is a senotherapeutic option for preventing the accumulation of senescent T cells in mice. Nat Commun. 2020;11(1):2482.
- Amor C, Feucht J, Leibold J, et al. Senolytic CAR T cells reverse senescence-associated pathologies. Nature. 2020;583(7814):127-132.
- Aghajanian H, Kimura T, Rurik JG, et al. Targeting cardiac fibrosis with engineered T cells. Nature. 2019;573(7774):430-433.
- Rurik JG, Tombácz I, Yadegari A, et al. CAR T cells produced in vivo to treat cardiac injury. Science. 2022;375(6576):91-96.
DECLARACIÓN DE TRANSPARENCIA
Los autores/as de este artículo declaran no tener ningún tipo de conflicto de intereses respecto a lo expuesto en el presente trabajo.
María Mittelbrunn
C/ Nicolás Cabrera, 1 · 28049 Madrid, España
Tlf.: +34 911 96 45 28 | E-Mail: mmittelbrunn@cbm.csic.es
Año 2022 · número 139 (02) · páginas 150 a 155
Enviado: 15.09.22
Revisado: 19.09.22
Aceptado: 24.09.22