

Resumen
La neuroplasticidad para procesar información táctil que genere una respuesta visual no es todavía bien conocida neurobiológicamente. No obstante, hay estudios que demuestran que el tacto pasivo permite percibir el objeto globalmente y en milisegundos. La percepción visual tiene lugar principalmente en nuestro cerebro y no en nuestros ojos. Este fenómeno se explica desde la neuroplasticidad crosmodal táctil, que consiste en la capacidad cerebral mediante la cual los individuos ciegos reorganizan y activan áreas cerebrales occipitales relacionadas con la visión a través del tacto, desde el córtex parietal primario o desde el tálamo hacia el lóbulo occipital. Es un proceso que se lleva a cabo mediante la experiencia de patrones de estimulación repetitiva táctil a través de las vías somatosensoriales durante largos periodos de tiempo.
La edad de comienzo de la ceguera, así como el momento en el que se inicia la estimulación táctil repetitiva son claves en el desarrollo de nuevas vías hacia el lóbulo occipital responsable de la visión. De hecho se sabe que los niños en los primeros años de vida generan los circuitos neuronales cerebrales mediante la experiencia temprana y la consolidación, tanto estructural como funcional, de las entradas sensoriales, que son a su vez mediadas por una reorganización de proteasas de la matriz extracelular, proceso que solo ocurre durante este período crítico.
La estimulación táctil pasiva diaria y sistemática permitirá una mayor conectividad en las áreas occipitales con una actividad neuronal alterada, como consecuencia de la pérdida de un sistema sensorial -en este caso la vista-. Dicha alteración conlleva, en gran parte, una falta de ¨poda¨ de las conexiones sobrantes no activas durante etapas del desarrollo cerebral en el propio lóbulo occipital o alteraciones en la via del tálamo al lóbulo occipital o ambas.
Abstract
The tactile neuroplasticity of the brain responsible for generating visual response yet remains to be known. However, studies show that, like vision, passive tactile sense enables the object to be perceived globally within milliseconds. The visual perception takes place mainly in our brain and not in our eyes. This can be explained by tactile crossmodal neuroplasticity consisting of the brain capacity that enables blind people to reorganize and activate occipital brain areas, related to vision, through touch, either from the primary parietal cortex or from the thalamus to the occipital lobe. This process is carried out through the experience of tactile repetitive stimulation patterns along the somatosensory pathways over longer periods of time.
The age of onset for blindness, as well as the time at which repetitive tactile stimulation may begin are key in achieving the development of new pathways towards the occipital lobe, which is responsible for vision. In fact, it is known that children in the first years of life generate brain neural circuits by early experience, as well as the structural and functional consolidation of sensory inputs which are mediated in turn by an extracellular matrix proteases reorganization. This process only occurs during such critical period.
Daily and systematic passive tactile stimulation will allow a greater connectivity in those occipital areas whose neuronal activity is altered as a result of a sensory system loss -in this case, sight- which largely entails a lack of pruning of the remaining non-active connections during certain brain development stages from within the occipital lobe and or alteration of the pathway from thalamus to occipital lobe or both.
Palabras clave: Neuroplasticidad; Visión táctil Tacto pasivo; Córtex occipital.
Keywords: Neuroplasticity; Tactile Vision; Passive Tactile Stimulation; Occipital Cortex.
INTRODUCCIÓN
La posibilidad de que los ciegos puedan ver se basa en el proceso neurobiológico de plasticidad cerebral, que consiste en la capacidad de las neuronas de comunicarse entre ellas, generar conexiones, establecer nuevas rutas cerebrales o reorganizar la estructura cerebral según la experiencia o la estimulación ambiental. Sin embargo, esta potencial habilidad de las neuronas táctiles para procesar información que genere una respuesta visual todavía no es bien conocida y está poco desarrollada por la neurociencia
El tacto es uno de los sentidos más importantes para nuestra supervivencia biológica: es el primer sistema de interacción con el medio ambiente, tanto en el aspecto de subsistencia como en el de interacción social. El tacto, además, nos sirve para reconocer texturas y formas de objetos o lectura de textos en braille a través del modo activo; esto es, utilizando nuestros dedos para explorar y percibir nuestro entorno, lo que conlleva mucho tiempo de análisis de la información. Este sistema de exploración tan lento -que va de las partes al todo- no permite que el cerebro pueda llegar a tener la sensación visual. En cambio, el tacto pasivo permite percibir el objeto globalmente y en milisegundos, al igual que la visión. Si esto es así, ¿se puede ver sin ojos?
Uno de los pioneros de la visión táctil, Bach y Rita (1), llegó a decir que la percepción visual tiene lugar principalmente en nuestro cerebro y no en nuestros ojos. De esta manera, si asumimos que “ver” es la capacidad de crear una representación mental de los objetos, su forma, propiedades, si esto es así, la posibilidad de que nuestros invidentes consigan ver es muy asequible y fácil de llevar a cabo.
Utilizando un modelo de estimulación táctil análogo al modelo visual, el cerebro podrá comprender de forma rápida, global, cognitiva y espacialmente integrada el entorno de forma muy parecida a como lo hace la visión mediante vías cerebrales similares.
La vía visual nace en la retina y llega a los núcleos geniculados laterales del tálamo y de ahí pasa directamente al córtex occipital primario, responsable de la visión, para completar el reconocimiento visual en áreas multimodales temporo-parieto-occipitales, mientras que la vía táctil parte de los receptores táctiles de la mano, llega a los núcleos sensoriales del tálamo y de ahí al córtex parietal primario, responsable de la sensación táctil, para terminar en áreas multimodales cerebrales. Ambas hacen un recorrido similar en el cerebro para llegar al reconocimiento de los objetos (Figura 1).
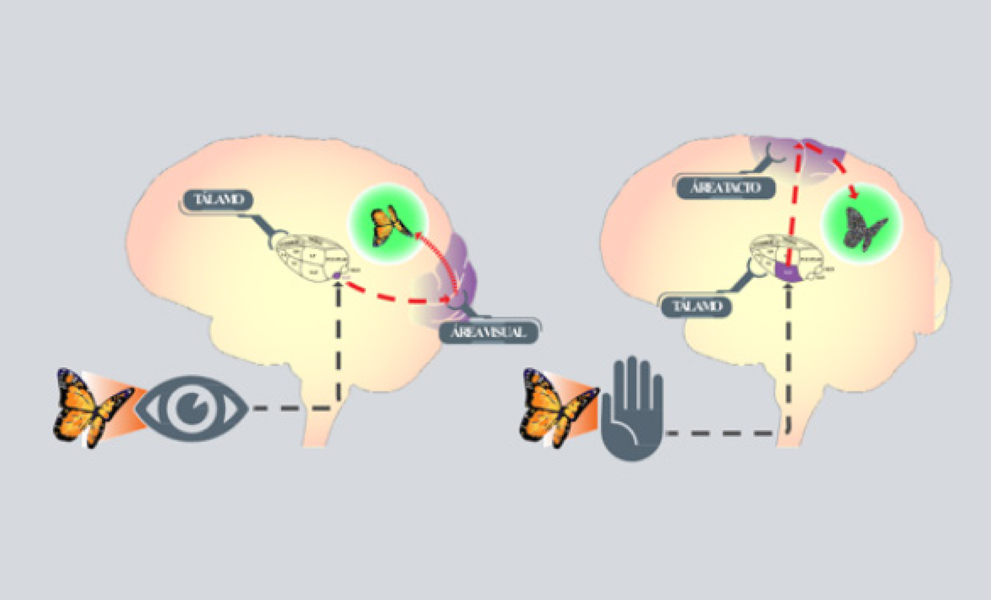
Con el propósito de conseguir visión táctil en sujetos ciegos necesitamos, por un lado, pasar del tacto activo al tacto pasivo mediante un sistema sustitutorio de estimulación táctil (SSET) (2), que evite el proceso exploratorio, lento, parcial y con movimiento de los dedos, y que permita captar las características espaciales del objeto percibido de forma global, simultánea y en milisegundos y por otro, un sistema de estimulación y entrenamiento táctil diario que posibilite una neuroplasticidad eficiente, estable y duradera, capaz de generar nuevas conexiones cerebrales que favorezca el “refuerzo de vías orgánicas preestablecidas, la formación de vías nuevas por ramificación y crecimiento progresivo de la arborización dendrítica y terminales nerviosas”, como decía Ramón y Cajal (3). La plasticidad neuronal se lleva a cabo mediante la experiencia de patrones de estimulación repetitiva táctil a través de las vías somatosensoriales. La actividad física regular y sistemática durante mucho tiempo estimula, tanto las conexiones inter- e intra-neuronal como el desarrollo de nuevas células nerviosas, sobre todo en el hipocampo (4,5).
Una de las ventanas de oportunidad de nuestros niños ciegos se da en los primeros años de vida, donde los circuitos neuronales en el cerebro están formados por la experiencia temprana. De hecho se sabe que la consolidación estructural y funcional de las entradas sensoriales están mediadas por una reorganización de proteasas de la matriz extracelular que ocurre solamente durante el período crítico (6-8).
El objetivo de la presente revisión es demostrar las posibilidades que ofrece la puesta en práctica de la estimulación táctil repetitiva pasiva durante varios meses, para generar plasticidad cortical, tanto en áreas occipitales responsables del reconocimiento viso-espacial en sujetos ciegos, como en áreas occipitales primarias responsables de la visión humana.
Estado del arte
Uno de los procesos neurobiológicos que puede ayudarnos a comprender cómo sería posible la visión a través del tacto es la «neuroplasticidad crosmodal», es decir, la capacidad cerebral mediante la cual los individuos ciegos reorganizan y activan áreas cerebrales occipitales relacionadas con la visión para procesar la información sensorial recibida a través del tacto (9-13). En realidad lo que hace el cerebro es desarrollar sistemas compensatorios de organización neurofuncional en las personas ciegas, de tal manera que sean capaces de integrar los estímulos táctiles en un proceso interactivo con el medio que llega a conseguir procesos sensoriales, perceptivos, cognitivos y emocionales similares a los que se llevan a cabo mediante la visión o la audición.
Si tenemos en cuenta esta perspectiva neurobiológica de la visión, parece menos paradójico preguntarse si el cerebro de una persona ciega puede ver, al menos si concebimos la visión como la capacidad de generar representaciones mentales internas que contienen detalles visuales. Aquí se hace necesario hablar de los «qualia visuales», que consisten en sensaciones ópticas, independientemente del origen de los estímulos. Para desarrollarlos en personas ciegas habrá que hacer uso de sistemas sustitutorios que lleven a cabo un proceso de estimulación táctil cercano o similar al neurobiológico de la visión. Las evidencias científicas demuestran la posibilidad de encontrar los sustratos neurobiológicos de los qualia visuales en ciegos mediante estímulos sensoriales auditivos o táctiles. En un estudio que hemos llevado a cabo, tras exponer a sujetos ciegos a estímulos táctiles de manera repetida y durante un largo período, se ha conseguido activar el lóbulo occipital al mismo tiempo que logrado una sensación subjetiva de qualia visual vinculada a la forma y orientación de estos estímulos hápticos después de un largo tiempo de entrenamiento táctil de líneas (14) (15) (Figura 2).

Lo peculiar de estos resultados es que los sujetos videntes, después de un entrenamiento táctil de varios meses de líneas, no consiguieron ver ni tener qualia visual; en cambio sí consiguieron reconocer las líneas en la palma de la mano. Una posible explicación a este hecho es que la estimulación diaria y continuada del sentido de la vista ha conseguido desarrollar grandes áreas cerebrales para la interpretación de los estímulos visuales, y dejado poco espacio cerebral para otras modalidades sensoriales como el tacto. El proceso visual genera una amplia red de conexiones asociadas con áreas visuales distribuidas en la parte posterior de nuestro cerebro, puesto que la información espacial es principalmente visual, y su automatización es tan alta que la implicación del tacto no tiene una respuesta eficiente a dicha red neuronal visual; por eso mismo se hace tan difícil que en una persona vidente se pueda llegar mediante neuroplasticidad a áreas visuales primarias que permitan una sensación visual. Sin embargo, en sujetos ciegos, sean tempranos o tardíos, se observa la capacidad de llegar a áreas occipitales mediante la estimulación táctil pasiva, aunque no debemos obviar que la edad influye notablemente en el grado de rapidez y desarrollo cerebral de las áreas occipitales que han recibido esta estimulación táctil, de tal forma que la activación occipital en el ciego temprano parece reflejar plasticidad crosmodal “pura”; por el contrario, observamos que en individuos ciegos tardíos la activación generada es occípito-temporal, como resultado, probablemente, de la mediación de imágenes visuales adquiridas antes de la ceguera (16-18) (Figura 3).
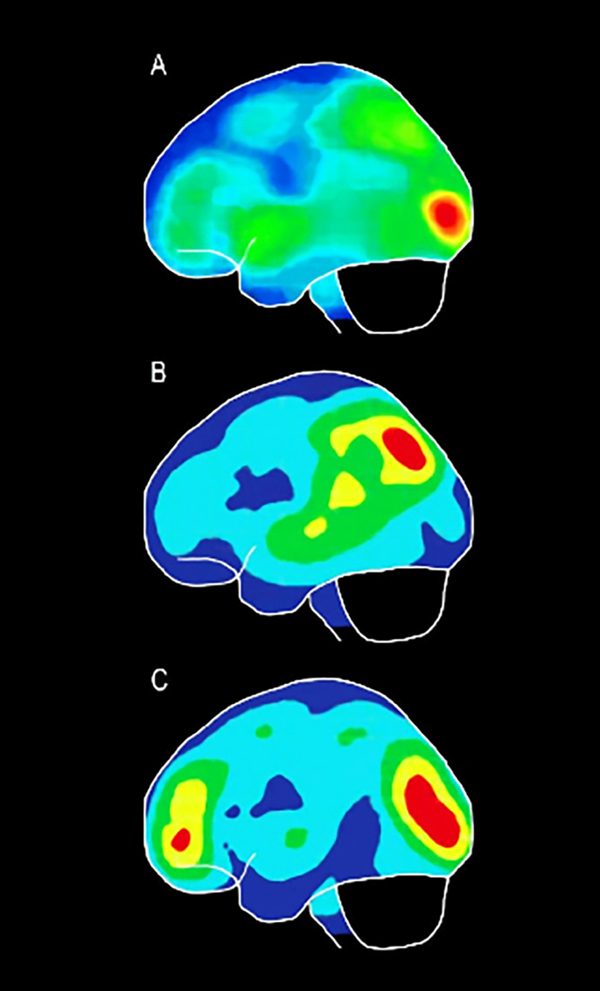
Nuestras investigaciones concluyen, además, que la estimulación táctil pasiva repetitiva a lo largo de periodos largos de entrenamiento consigue una neuroplasticidad estable y permanente en las áreas occipitales primarias de sujetos ciegos, independientemente de la edad de inicio de la ceguera, y que en algunos casos conlleva qualia visual. Sin embargo, otros investigadores han demostrado que, también en períodos de estimulación de tiempo cortos, tienen lugar procesos de reorganización cerebral, tanto es así que existe incluso evidencia de que los fenómenos de reorganización crosmodal pueden inducirse con muy pocos días, horas o tan solo minutos de inactividad visual y/o actividad táctil (19,20).
Este proceso se debe principalmente a la neuroplasticidad crosmodal, que a través de la estimulación táctil es capaz de activar áreas cerebrales visuales. Lo que hace el cerebro, en definitiva, es otorgar un output (p. ej. visual) en una modalidad sensorial distinta de la modalidad de input -en nuestro caso, táctil. Otros investigadores (21,22) comprobaron que la lectura braille indujo en sujetos ciegos tempranos respuestas bilaterales en la corteza visual, mientras que la mayoría de los sujetos ciegos tardíos mostraron respuestas solamente ipsilaterales en la corteza occipital inferior; asimismo, encontraron, durante la estimulación vibrotáctil, mayor actividad de la corteza visual en el grupo de ciegos de inicio temprano que en el de inicio tardío, y una disminución de la actividad en cortezas visuales asociativas cuanto más tardía es la aparición de la ceguera (23-26). Un estudio llevado a cabo por Voss y col. (27) demostró que los ciegos tardíos mostraban mayor actividad en áreas visuales ventrales de la corteza occípito-temporal, que los ciegos tempranos, que presentaban una activación mayor en estriado y corteza extraestriada. A la vista de estos datos podríamos decir que, mientras que la activación occipital en el ciego temprano parece reflejar plasticidad crosmodal “pura”, la activación occípito-temporal en individuos ciegos tardíos puede ser el resultado de la mediación de imágenes visuales adquiridas antes de la ceguera. Sin obviar lo dicho, Gougoux y col. (28) encontraron que también los estímulos auditivos indujeron un patrón similar de activación en la corteza occipital en ambos grupos de ciegos, tardíos y tempranos, aunque la activación cerebral fue más fuerte en el surco temporal superior izquierdo del grupo de ciegos congénitos frente al grupo de ciegos tardíos y videntes.
CONCLUSIONES
Mediante el entrenamiento táctil pasivo diario, tal y como hemos visto con los niños en los estudios presentados, conseguiremos que personas ciegas desarrollen neuroplasticidad de áreas sensoriales visuales, multimodales y de integración y su consiguiente capacidad de procesar, identificar y reconocer la información sensorial táctil pasiva recibida. La estimulación táctil pasiva diaria y sistemática permitirá una mayor conectividad en áreas occipitales que se encuentran con una actividad neuronal alterada, como consecuencia de la pérdida de un sistema sensorial (la vista) que conlleva, en gran parte, una falta de poda de las conexiones sobrantes no activas durante etapas del desarrollo cerebral.
A la vista de estos datos, y a pesar de que todavía no está claro el proceso neurofuncional con detalle, podríamos aventurarnos a decir que la aplicación de estímulos táctiles repetitivos pasivos activan áreas somatosensoriales primarias visuales y, a medida que avanza la estimulación táctil, se activan estructuras multimodales capaces de reconocer e identificar los estímulos espaciales, para acabar, al menos en algunos sujetos, en áreas visuales occipitales, lo que ofrece la sensación subjetiva de qualia visual. Los resultados de otras investigaciones, en cambio, favorecen más la plasticidad de la vía tálamo-occipital como responsable del proceso neurofuncional de la visión, mientras que otros establecen una vía crosmodal táctil más lenta que llega a las áreas occipitales pero sin la capacidad de generar visión (13). Una posible explicación es que mediante la visión retina-tálamo-córtex occipital las respuestas son predominantemente excitatorias, mientras que en la vía tacto-córtex-parietal-córtex occipital, vía neuroplasticidad crosmodal, las respuestas son predominantemente inhibitorias (29). Este proceso podría deberse a que no haya una actividad que siga un patrón definido lo suficientemente estable y robusto que consiga la modificación de la función del canal iónico en el axón, las dendritas y el cuerpo celular capaz de producir los cambios específicos necesarios para la plasticidad sináptica de los potenciales postsinápticos excitadores o inhibidores (30,31).
Dentro del objetivo de acercar al sujeto invidente el modelo visual del individuo vidente, no nos cabe la menor duda de que nuestros avances científicos conseguirán que las personas ciegas, en un futuro, puedan identificar formas, dimensiones, colores, no solo de forma verbal, como hasta ahora lo hacen, sino también de forma perceptiva. Futuros estudios podrán ayudar a entender los procesos neurofisiológicos subyacentes a los fenómenos visuales y al reconocimiento espacial mediante estimulación táctil pasiva.
BIBLIOGRAFÍA
- Bach y Rita, P., Brain mechanisms in sensory substitution. Academic Press. 1972.
- Bach y Rita, P., Colliins, C.C., Saunders, F.A., Whi- te, B., Scadden, L. Vision substitution by tactile image projection. 1969 Nature; 221: 963–964.
- Ramón y Cajal, S. Textura del sistema nervioso del hombre y de los vertebrados. Gobierno de Aragón, Zaragoza, 1ª edición (edición facsímil), 1889.
- Van Praag H., Christie B.R., Sejnowski T.J., Gage F.H. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc Nat Ac Science 1996; 96(9): 13427-13431.
- Gheusi G., Rochefort C. Neurogenesis in the adult brain. Functional consequences. J Soc. Biol, 2002, 196(1): 67-76.
- Shiosaka, S., Yoshida, S. Synaptic microenvironments, structural plasticity, adhesion molecules, proteases and their inhibitors. Neurosci. Res . 2000; 37: 85–89.
- Ismail FY, Fatemi A, Johnston MV. Cerebral plasticity: Windows of opportunity in the developing brain. Eur J Paediatr Neurol. 2017, 21(1):23-48.
- Henschke JU, Oelschlegel AM, Angenstein F, et al. Early sensory experience influences the development of multisensorial thalamocortical and intracortical connections of primary sensory cortices. Brain Struct Funct. 2018;223(3):1165-1190
- Merabet, LB., Hamilton, R., Schlaug,G., et al. Rapid and reversible recruitment of early visual cortex for touch. PLoS ONE, 2008, 3 (8):e3046.
- Sathian, K. y Stilla.R. Cross-modal plasticity of tactile perception in blindness. Rest Neurol Neurosci, 2010, 28 (2):271–281.
- Stilla, R., Hanna, R., Hu, X., Mariola, E., Deshpande,G., Sathian, K. Neural processing underlying tactile microspatial discrimination in the blind: A functional magnetic resonance imaging study. J Vision 2008, 8 (10):1–19.
- Henschke, J.U., Noesselt, T., Scheich, H., Budinger, E. Possible anatomical pathways for short-latency multisensory integration processes in primary sensory cortices. Brain Struct Funct 2015;220:955–977.
- Muller, F., Niso, G., Samiee, S., Ptito, M., Baillet, S., Kupers, R. A thalamocortical pathway for fast rerouting of tactile information to occipital cortex in congenital blindness. Nat Commun 2019;10:1-9
- Ortiz T., Poch J., Santos J.M., et al.
Occipital enduring neuroplasticity induced by long-term repetitive tactile stimulation: a case report, Neurocase 2014;20(3):273-282. - Ortiz T., Santos JM., Generación de experiencias visuales en ciegos mediante estimulación táctil repetitiva, Ciencia Cognitiva 2012;6(1): 9-12.
- Sadato N,Okada T, Honda M, Yonekura Y. Critical period for cross-modal plasticity in blind humans: A functional MRI study. Neuroimage, 2002; 16 (2):389–400.
- Sani L, Ricciardi E, Gentili C et al.. Effects of visual experience on the human MTþ functional connectivity networks: An fMRI study of motion perception in sighted and congenitally blind individuals. Front Sys Neurosci 2010;4:159
- Collignon O, Dormal G, Albouy G, et al. Impact of blindness onset on the functional organization and the connectivity of the occipital cortex. Brain 2013;136:2769–2783
- Boroojerdi B., Bushara K. O., Corwell B. et al. Enhanced excitability of the human visual cortex induced by short-term light deprivation. Cerebral Cortex. 2002;10 (5):529–534.
- Facchini, S. Y Aglioti, SM. Short term light deprivation increases tactile spatial acuity in humans. Neurology, 2003; 60 (12):1998–1999.
- Burton H., Snyder A.Z., Contouro T.E. et al. Adaptive changes in early and late blind: An fMRI study of Braille reading. J Neurophysiol 2002; 87 (1):589–607.
- Burton, H. Visual cortex activity in early and late blind people. J Neurosc, 2003; 23 (10):4005–4011.
- Burton H, Sinclair RJ, McLaren DG. Cortical activity to vibrotactile stimulation: An fMRI study in blind and sighted individuals. Hum Brain Mapping, 2004; 23 (4):210–228
- Burton, H., McLaren, DG, Sinclair, RJ. Reading embossed capital letters: An fMRI study in blind and sighted individuals. Hum Brain Mapping, 2006; 27 (4):325–339.
- Burton, H., D. G. McLaren. Visual cortex activation in late-onset, Braille naive blind individuals: An fMRI study during semantic and phonological tasks with heard words. Neurosci Letters, 2006; 392 (1–2):38–42.
- Leo A, Bernardi G, Handjaras G, et al. Increased BOLD variability in the parietal cortex and enhanced parieto- occipital connectivity during tactile perception in congenitally blind individuals. Neural Plast 2012;2012:720278
- Voss, P., Gougoux, F., Zatorre,RJ., et al. Differential occipital responses in early- and late-blind individuals during a sound-source discrimination task. Neuroimage, 2008, 40 (2):746–758.
- Gougoux F., Belin P., Voss P., et al. Voice perception in blind persons: A functional magnetic resonance imaging study. Neurophysiol 2009; 47 (13):2967–2974.
- Nasconcelos N., Pantojaf J., Belchiora H. et al. Cross-modal responses in the primary visual cortex encode complex objects and correlate with tactile discrimination. PNAS 2011;108;(37): 15408-15413
- Karlen SJ, Kahn DM, Krubitzer L. Early blindness results in abnormal corticocortical and Thalocortical connections. Neuroscience 2006;142(3): 843-858
- Kemenes I., Straub V.A., Nikitin E.S. et al. Role of Delayed Nonsynaptic Neuronal Plasticity in Long-Term Associative Memory. Curr Biol
DECLARACIÓN DE TRANSPARENCIA
El autor/a de este artículo declara no tener ningún tipo de conflicto de intereses respecto a lo expuesto en la presente revisión.
ranm tv
Tomás Ortiz Alonso
Departamento de Medicina Legal, Psiquiatría y Patología
Facultad de Medicina.Universidad Complutense de Madrid
Avenida Complutense, s/n · 28040 Madrid
Tlf.: +34 91 39 41 497 | E-Mail: tortiz@ucm.es
Año 2020 · número 137 (01) · páginas 22 a 26
Enviado*: 11.02.20
Revisado: 18.02.20
Aceptado: 14.03.20
* Fecha de lectura en la RANM