

Resumen
En 1859 Charles Darwin en su “The Origin of Species” formuló tres hechos que explican el principio de la evolución: la presencia de un antepasado común, la existencia de mutaciones genéticas ventajosas para la supervivencia y eliminación de las desfavorables o Selección Natural. El desarrollo embriológico del corazón de los mamíferos mejora la necesidad de captación del oxígeno exterior, su transporte y liberación en los tejidos, que existe en todos los seres vivos pluricelulares desde los más primitivos y posibilita la función de todos los órganos y sistemas. En los invertebrados, como insectos, artrópodos y moluscos, el sistema circulatorio abierto alcanza un desarrollo similar al de los estadios 10-13 de Carnegie humanos: tubos endocárdicos, tubo recto o primeras fases del Asa bulbo ventricular. En los vertebrados, el sistema cerrado de los peces, desarrolla corazones de tres y cuatro cámaras equivalentes a la fase de torsión del asa en los mamíferos, o quinta semana de desarrollo embrionario, apareciendo en los reptiles un modelo más próximo al de los mamíferos con diferencias como la partición incompleta de los ventrículos y el desarrollo de dos troncos aórticos en los cocodrilos. Aves y mamíferos alcanzan un grado de desarrollo embrionario de su corazón similar, con las cuatro cámaras y el tronco cono septados, que separan la circulación sistémica y pulmonar. Todo el desarrollo embriológico del corazón humano se completa entre el día 20 y 60 de gestación, concluyendo en la fase fetal su crecimiento junto al resto del organismo.
Abstract
In 1859 Charles Darwin in his “The Origin of Species” formulated three facts that explain the principle of evolution: the presence of a common ancestor, the existence of advantageous genetic mutations for survival and the elimination of unfavorable ones or Natural Selection. The embryological development of the mammalian heart improves the need for external oxygen uptake, transport and release in the tissues, which exists in all multicellular living beings from the most primitive and enables the function of all organs and systems. In invertebrates, such as insects, arthropods and molluscs, the open circulatory system reaches a development similar to that of human Carnegie stages 10-13: endocardial tubes, straight tube, or early stages of the bulboventricular loop. In vertebrates, the closed system of fish develops hearts with three and four chambers equivalent to the torsion phase of the loop in mammals, or the fifth week of human embryonic development, with a model closer to that of mammals appearing in reptiles with differences such as the incomplete septation of the ventricles and the development of two aortic trunks in crocodilians. Birds and mammals have reached a similar degree of embryonic development of their hearts, with four septate chambers, the same as the Cono Truncus one, and which separate the systemic and pulmonary circulation. All the embryological development of the human heart is completed between days 20 and 60 of gestation, concluding in the fetal phase its growth together with the rest of the organism.
Palabras clave: Asa; Bulbo Ventricular; Sistema circulatorio abierto; Sistema circulatorio cerrado; Corazón bicameral; Corazón tricameral; Corazón cuatro cámaras.
Keywords: Bulboventricular Loop; Open circulatory system; Closed circulatory system; Bicameral heart; 3 chambered heart; Heart four chambers.
INTRODUCCIÓN
En 1859 Charles Darwin publicó “The Origin of Species”. Nos permitió entender que las especies no son permanentes, ni inmutables y que no fueron creadas. Hace más de tres mil millones de años (“mda”) comenzó la vida unicelular. Durante centenares de “mda” han ido apareciendo a partir de un antepasado común los 40 Filos de especies animales que conocemos(1). Las mutaciones genéticas produjeron los cambios que favorecieron la supervivencia, mientras que mutaciones deletéreas permiten eliminar al individuo que la sufre (Selección Natural)(2).
LA EVOLUCIÓN EMBRIOLÓGICA
Toda la evolución, según Darwin, se realiza de forma gradual y continua, sin cambios abruptos, y se refleja en el desarrollo embriológico de un órgano que muestra que en antepasados remotos, ese órgano alcanzó un desarrollo completo que aparece reflejado en algún estadio del desarrollo embriológico en especies superiores(1).
DESARROLLO EMBRIOLÓGICO DEL CORAZÓN HUMANO
El desarrollo embrionario del corazón, se debe asociar con clasificaciones de estadificación general de los embriones (3) como los 23 estadios de Carnegie que se completan en 56-60 días, (4). Añadiendo a las características anatómicas el tiempo de desarrollo y el tamaño del embrión en cada etapa (5).
En las dos primeras semanas de desarrollo embrionario no hay vestigios de corazón ni grandes vasos. A partir del día 20, (estadio 9 de Carnegie) el embrión mide 1,5 a 2,5 mm se inicia el desarrollo embrionario del corazón. Se esbozan dos primordios cardiacos, a la derecha e izquierda de la línea neural primitiva del disco embrionario, crecen uniéndose por su parte cefálica, conformando un cuerpo cardiogénico en forma de herradura. El día 21, los primordios, forman dos tubos endocárdicos individuales, a ambos lados de cresta neural.
Estadio 10 de Carnegie (días 22-23; 3 mm). Los dos tubos se fusionan en el centro de la cavidad celomática pericárdica, dando lugar a un tubo único, el Tubo Recto (TR), con material genético de ambos lados. Es el vestigio del Ventrículo Primitivo (VP) o futuro ventrículo izquierdo (VI). Conecta caudalmente con las aurículas primitivas derecha e izquierda que permanecen individualizadas hasta el corazón desarrollado. Se separan del VP, por los surcos atrioventriculares (AV) (6,7) (Figura 1).

Craneal al Tubo Recto aparece el Bulbus Cordis (BC), futuro ventrículo derecho (VD). Se separa del VP por el surco inter bulbo ventricular (BV). Se seguirá incorporando cefálicamente el Cono o cámara de salida ventricular, que más adelante conecta con el Truncus que dará origen al tronco principal de la aorta y pulmonar, que se unirán a los arcos branquiales o aórticos de los que se originan el cayado aórtico, las ramas de la arteria pulmonar y distintas arterias sistémicas supraaórticas (Figura 1).
Día 25 (estadio 12 de Carnegie. Días 22 y 28; 3-5 mm) el TR inicia una flexión y torsión dentro de la cavidad celómica pericárdica. Profundiza el surco inter BV izquierdo y va desapareciendo el derecho. Se forma así, el Asa BV a la derecha. En un proceso ininterrumpido el Asa BV, se torsiona hacia delante; el BC se traslada hacia abajo, a la derecha y por delante del VP y va apareciendo el cono y el tronco. El VP va ascendiendo por detrás (y con él las aurículas) del BC (Figura 1).
Las aurículas están conectadas con el VP y este con el BC que cefálicamente se continúa con el tronco cono (TC) o cámara de salida del BC. Las posibles futuras anomalías congénitas en el corazón desarrollado es más fácil que sean por detención que por sobrepasar el desarrollo normal. Por eso diagnosticamos doble cámara de entrada de ventrículo izquierdo (VI) (o ventrículo único) y doble cámara de salida del ventrículo derecho (VD), y es una rareza una doble salida del VI o doble entrada del VD.
Las aurículas se desplazan de la posición más caudal a una posición dorsal y craneal respecto al VP. Delante de las aurículas se apoya el TC, que comprime ligeramente la aurícula. En ese punto comienza a desarrollarse el septum primum que dividirá las aurículas. Por detrás se forma un pequeño receso, el seno venoso, en el que desembocan las venas onfalomesentéricas (umbilicales), vitelinas y cardinales y en el corazón desarrollado en el cuerno derecho del Seno Venoso lo harán las Venas Cavas y en el izquierdo las venas pulmonares.
Simultáneamente comienza la tabicación de las aurículas, de los ventrículos, del tronco cono de salida, y la delineación de las válvulas aurículo-ventriculares (AV) y las sigmoideas. Aparecen los cojines endocárdicos AV dorsal y ventral, que dividirán el canal AV cuando se fusionen en orificio derecho e izquierdo. Se ensanchará a la derecha, lo que incorporará la cámara de entrada al futuro VD El cono posterior se incorporá al VP para proporcionar cámara de salida al futuro VI.(Figura 2)
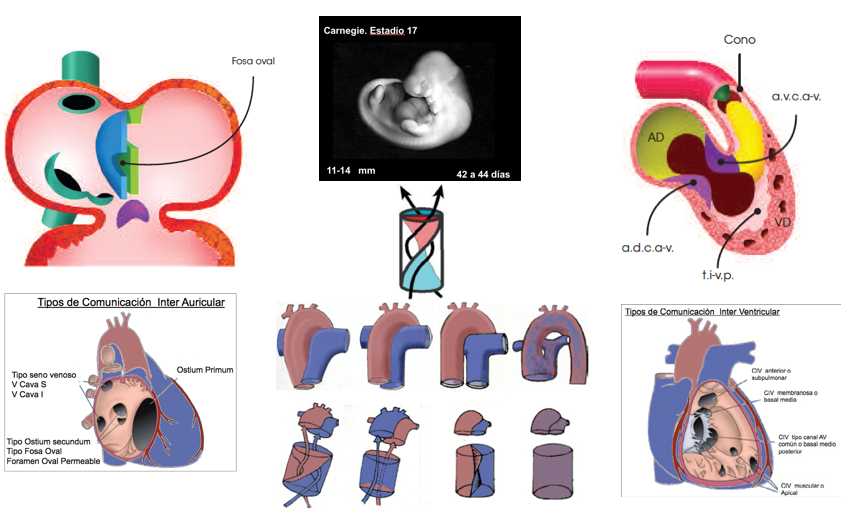
En el interior del surco BV izquierdo se forma una cresta prominente denominada espolón BV que separa las paredes contiguas del BC y VP y que llegará a reabsorberse. En el interior de la punta cardiaca entre el VP y el BC comienza a desarrollarse el septo o tabique interventricular (IV) primitivo (TIVP). Entre el TIVP y el espolón BV se forma el orificio o comunicación interventricular (CIV) primitiva, que conforme vaya desarrollándose el septo llegará a desaparecer, persistiendo al final del desarrollo embrionario solo la CIV subaórtica membranosa, que se cerrará con material genético de varios orígenes del tabique IV.
Final de la cuarta semana, (Estadio 13 de Carnegie días 28 a 32. 4-6 mm) ya están formadas las cuatro cavidades cardíacas primitivas y establecida la circulación embrionaria y extraembrionaria. Está completándose el septum interauricular (IA), y comienzan a excavarse las bolsas trabeculadas a ambos lados del TIVP, que crecerá hasta encontrarse con los cojines endocárdicos cerrando el ostium primum y las crestas conales que acabarán cerrando la CIV primitiva. La CIV membranosa subaórtica, será la última comunicación en cerrarse.(Figura 2)
Días 32 y 33 ( 7 mm) (Estadio 14 de Carnegie) , septación del TC por las crestas tronco conales, que giran 180º y dividen el TC en cámara de salida de los ventrículos y troncos arteriales de aorta y pulmonar. El cono anterior conecta con el tronco posterior y este con el sexto arco aórtico que da origen a las ramas de la arteria pulmonar. El cono posterior, con el giro de las CTC conecta con el tronco anterior, y este con el 4ª arco aórtico que origina el cayado aórtico.(Figura 2).
El VD adquiere la cámara de entrada al expanderse el canal AV hacia el BC-VD cuando se inicia la división de los septos, IA e IV. La septación conal, incorpora el cono posterior al VP, o cámara de salida del VI. En la semana 7 Días 51-53. (18 – 22 mm) (estadio 20) se ha completado el desarrollo embrionario del corazón.
EVOLUCIÓN FILOGENÉTICA DEL CORAZÓN
Hace tres mil millones de años (“mda”), 1500 después de que se formara la Tierra, comenzó la vida con la aparición de células eucariotas o nucleadas unicelulares. Hace unos 800 “mda”, aparecieron animales pluricelulares con simetría radial(8). A partir de estos animales, en el transcurso de centenares de “mda” fueron apareciendo nuevas especies de animales invertebrados y vertebrados. Muchas de ellas se han extinguido.
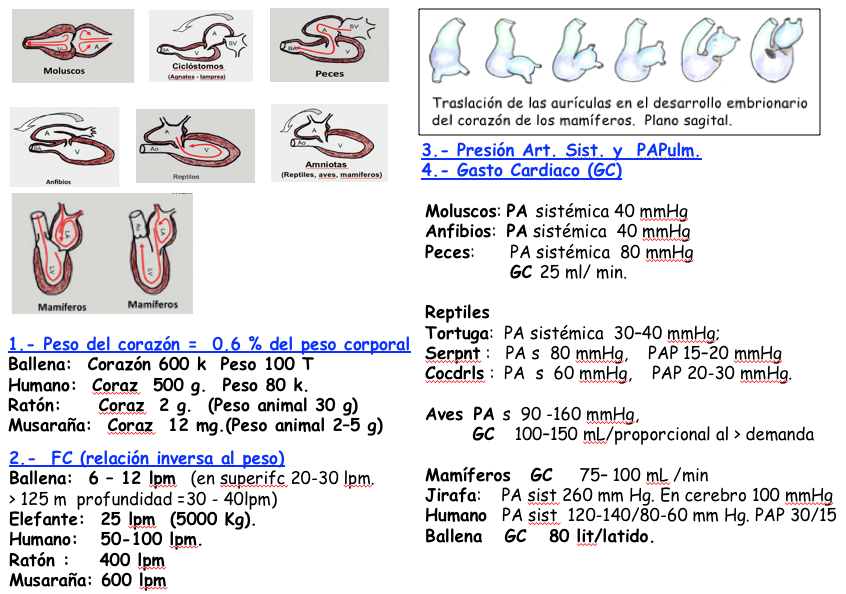
Todos los animales precisan de un sistema circulatorio capaz de transportar oxígeno y nutrientes. Necesita un fluido circulante (sangre o hemolinfa), vasos sanguíneos y uno o varios corazones. Según cómo circule el fluido el sistema puede ser abierto o cerrado. Sistema circulatorio abierto: lo tienen muchos invertebrados. El corazón impulsa la hemolinfa por arterias al espacio cavitario o hemocele, donde se intercambian nutrientes, gases y desechos y retorna al corazón por venas. El sistema circulatorio cerrado lo poseen anélidos, cefalópodos y vertebrados. El corazón impulsa la sangre por el sistema arterio-venoso.
CORAZÓN EN LOS INVERTEBRADOS
Entre los invertebrados que no tienen simetría radial se encuentran los Poríferos, como las esponjas. Aparecieron hace 640 “mda” carecen de verdadero sistema circulatorio y realizan el intercambio de nutrientes, oxígeno y desechos directamente con el medio acuático en el que viven y con sus células, mediante difusión. Carecen también de sistema respiratorio(9).
Los platelmintos (gusanos planos), aparecieron hace 540-500 “mda”. Son precursores de los gusanos. Primeros representantes de animales con simetría bilateral (10). No tienen todavía un sistema circulatorio con órganos específicos.
Los Anélidos como las Lombrices de tierra, aparecieron hace 650 a 570 “mda”. Tienen ya un sistema circulatorio cerrado con un gran vaso dorsal y otro ventral, comunicados por vasos transversales. Todos los vasos se contraen rítmicamente. Los vasos transversales anteriores son los arcos aórticos y actúan como pequeños corazones (11) Respiran por difusión a través de la piel.
Su sistema circulatorio puede corresponder con el desarrollo de los dos tubos endocárdicos bilaterales en los embriones de los mamíferos.
Insectos
Los primeros insectos aparecieron en la Tierra hace 480 “mda”, Su sistema circulatorio es abierto. Con un vaso dorsal con dos partes una caudal hacia el hemocele abdominal, en medio el corazón capaz de contraerse muy rápidamente y enviar la hemolinfa por el vaso cefálico a la cavidad o hemocele craneal. La respiración la realizan directamente por un conjunto de tubos abiertos al exterior por unos orificios, espiráculos, en tórax y abdomen. Presentan también corazones pulsátiles accesorios independientes para ayudar a la circulación en la raíz de las alas (moscas) o extremidades (langostas)(12).
Los crustáceos aparecieron hace 540 “mda”. Presentan varios tipos de sistemas cardiovasculares. Desde conductos peristálticos primitivos en pequeños organismos del plancton a un sistema circulatorio pulsátil complejo con hemocianina como vehículo del oxígeno, en decápodos (cangrejos, langostas) que poseen un corazón situado en el tórax, dentro de un pericardio rígido al que se fijan por ligamentos elásticos, que le facilitan la diástóle. La sangre venosa llena el interior del pericardio y en diástole ingresa al ventrículo a través de canales en la pared del miocardio. En sístole cierra estos orificios y eyecta la sangre a las aortas anterior y posterior. Las branquias, situadas en el territorio venoso, hacen que la sangre que recibe el corazón sea sangre arterial. Como los mamíferos, poseen un sistema neuro hormonal para el control de la frecuencia y el volumen latido(12).
Moluscos
Aparecieron hace 530 “mda”. Existen unas 100.000 especies y unas 30.000 se han extinguido. Salvo en los cefalópodos, el sistema circulatorio es abierto. El corazón, según la especie, tiene 2 ó 3 cavidades, un ventrículo y una o dos aurículas. Impulsa la hemolinfa por arterias hasta espacios o senos abiertos de donde la recogen las venas, en su trayecto se oxigena en las branquias y vuelve así al corazón.
Los cefalópodos (pulpos, calamares, sepias) tienen un sistema circulatorio cerrado. Presentan dos corazones branquiales que impulsan la hemolinfa a las branquias, donde se oxigena antes de pasar a las aurículas (12).
Ciclóstomos, (Peces primitivos)
Los Ciclóstomos (boca circular), aparecieron hace 510 “mda”. Los representantes actuales son los Agnatos (sin mandíbula) como las Lampreas. E sistema circulatorio es cerrado con un Corazón tricameral (Seno venoso, aurícula y ventrículo). Cuando se ha desarrollado aparece plegado, con una flexión en S en el plano sagital y en U en el transversal. El Seno venoso y la aurícula pasan a posición dorsal e izquierda; el ventrículo y BC (que puede desaparecer), pasan a la derecha. Arterias, capilares y venas completan el sistema CV. Desde el punto de vista evolutivo corresponde con la etapa de ASA BV de los mamíferos.
Peces
Los primeros peces con mandíbulas (gnatóstomos) aparecieron hace 485 “mda”. Tienen un sistema circulatorio con un corazón bicameral, aurícula y ventrículo. Este envía la sangre a las branquias donde los capilares la envían al resto del organismo y luego regresa al corazón. Fiilogenéticamente este corazón es similar a la fase embrionaria en los mamíferos de TR tras la fusión de los tubos endocárdicos.
Anfibios
Sapos, ranas y tritones evolucionaron hace 365 “mda” de su ancestro sarcopterigio (peces de aletas carnosas). Durante más de 100 “mda” fueron los vertebrados dominantes, hasta que aparecieron los reptiles. Su sistema circulatorio es un doble circuito a diferencia del circuito simple de los peces. Este sistema doble evolucionó para poder aportar un suministro adecuado de oxigeno a los tejidos mediante la separación de la sangre en el pulmón para ser oxigenada y pasar al circuito sistémico.
El corazón de los anfibios es una modificación del corazón rectilíneo de los peces (senovenoso, atrio, ventrículo y cono arterioso). Los anfibios poseen un corazón tricameral Seno venoso–AD , AI, ventrículo, y cono arterioso. La sangre de los pulmones entra por vena pulmonar a la AI, de ahí al VI, y se eyecta al organismo a través del cono arterioso (parte proximal de Ao ventral). El cono arterioso posee una válvula espiral musculosa, con una parte anterior y otra posterior. El ventrículo bombea sangre sistémica hacia los pulmones y sangre oxigenada hacia el resto del organismo a través de varias arterias y arcos arteriosos.
En relación al corazón de los mamíferos, el corazón de los anfibios podría corresponder a la fase post asa bulbo ventricular en que se ha comenzado a dividir el septo interventricular y también han aparecido las crestas TC. Clínicamente este tipo de corazón correspondería con un ventrículo único (doble cámara de entrada de VI) con un tronco común, difícilmente viable.
Reptiles
Aparecen hace unos 270 “mda”, a partir de un Batracio, el Eryops que medía 1,50 m de longitud. Los reptiles al pasar del medio acuático al terrestre, necesitaron desarrollar pulmones y circulación pulmonar. No existe un modelo único, sino que el corazón de los reptiles está condicionado por la anatomía del tipo de reptil y su fisiología condicionada por el medio en que desarrolla su vida, acuáticos, terrestres o combinados. Un “típico” corazón reptiliano de tortugas y saurios tiene tres cámaras, dos aurículas (derecha e izquierda) y un ventrículo.
CORAZÓN EN LOS VERTEBRADOS
Los vertebrados salieron del medio acuático al terrestre hace 350 “mda”. Tuvieron que respirar aire y hacer frente a las fuerzas gravitacionales, características que explican la amplia variedad de morfologías del corazón entre los vertebrados. Desarrollaron un corazón multicámara con un sistema vascular cerrado con capilares, recubierto por endotelio, sangre con células características, eritrocitos con hemoglobina, leucocitos y células y factores de coagulación.
Las Aves derivan de los saurios terópodos, al final del Jurásico, hace unos 150 “mda”. Las aves poseen un corazón de cuatro cámaras y un sistema arteriovenoso con circulación pulmonar y sistémica. A diferencia del sistema de los mamíferos en las aves los eritrocitos son nucleados.
Mamíferos
Los primeros mamíferos aparecieron hace 205 “mda” y desarrollaron el corazón con cuatro cámaras, característico también de los humanos que hemos descrito en su desarrollo embriológico.
En conclusión todos los grupos animales, desde los más primitivos, desarrollaron algunas características en sus sistemas cardio vasculares, que fueron ventajosas para su supervivencia y la aparición a lo largo de millones de años, de nuevas especies que progresivamente permitieron que se alcanzara el desarrollo del sistema cardiovascular de los mamíferos culminado en el humano, que permitió el desarrollo de un sistema nervioso central inteligente gracias a mantener una circulación sistémica y pulmonar y unas constantes cardiovasculares como la Tensión arterial y el Gasto cardiaco. (Figura 3).
BIOGRAFÍA
- Charles Darwin. The origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. John Murray, Albemarle street. London.1859.
- Sánchez Ron J.M. Charles Darwin (1809-18829) Su obra y su mundo. En M Charles Darwin: Una vida y una visión del mundo. Sánchez Ron J.M (editor). Instituto de España, Madrid. 2009. 9-74
- Mall F.P. On stages in the development of human embryos from 2 to 25 mm long. Anat. Anz., 1914; 46: 78–84.
- O’Rahilly R. Early human development and the chief sources of information on staged human embryos. Europ. J. Obstet. Gynecol Reprod Biol. 1979; 9: 273-280.
- Jatsu Azkue J. External surface anatomy of the postfolding human embryo: Computer-aided, three-dimensional reconstruction of printable digital specimens J Anat. 2021; 239: 1438–1451.
- Berrazueta Fernández J.R. Berrazueta Sánchez de Vega A, Gutiérrez Escalada B, Viadero Ubierna M. Recuerdo embriológico del corazón y su alteración en las cardiopatías congénitas. En Libro de Cardiología de Valdecilla. Edit. J R Berrazueta; Edit asociado JA Vázquez de Prada. Edit Grupo Teiba. Guarnizo Cantabria. 2017. 521-534.
- Gómez-Gómez M, Danglot-Banck,C, Santamaría-Díaz H, Riera-Kinkel C. Desarrollo embriológico y evolución anatomofisiológica del corazón (Primera Parte). Rev Mex Pediat 2012; 79 : 92-101.
- Laos R, Benner S. El origen de la vida y la primera molécula capaz de replicarse a sí misma. Rev Química PUCP, 2014, 28: 24-33.
- Gold DA Grabenstatter J, Mendoza A, Riesgo A, Ruiz Trillo I, Summons RE. Sterol and genomic analyses validate the sponge biomarker hypothesis PNAS 2016; 113: 2684-2689.
- Ruiz Trillo I, Riutort M, Littlewood DTL, Herniou EA, Baguña J. Acoel flatworms: earliest extant bilaterian Metazoans, not members of Platyhelminthes. Science 1999; 283(5409):1919-23. doi: 10.1126/science.283.5409.1919.
- Nanglu K, Caron JB. A New Burgess Shale Polychaete and the Origin of the Annelid Head Revisited. Current Biology 2018; 28: 319-326. https://doi.org/10.1016/j.cub.2017.12.019
- Bettex DA, Prêtre R, Chassot PG. Is our heart a well-designed pump?. The heart along animal evolution. Eur Heart J. 2014; 35: 2322–2332. doi:10.1093/eurheartj/ehu222
DECLARACIÓN DE TRANSPARENCIA
El autor/a de este artículo declara no tener ningún tipo de conflicto de intereses respecto a lo expuesto en el presente trabajo.
ranm tv
José Ramón de Berrazueta Fernández
Real Academia Nacional de Medicina de España
C/ Arrieta, 12 · 28013 Madrid
Tlf.:+34 91 547 03 18 | E-Mail: joseramon@berrazueta.com
Año 2022 · número 139 (01) · páginas 16 a 22
Enviado*: 22.03.22
Revisado: 26.03.22
Aceptado: 30.03.22
* Fecha de lectura en la RANM