Resumen
Hiper-inflamación, hiper-coagulación y daño endotelial son hallazgos principales en el Covid-19 agudo o persistente. La proteína S del SARS-COV-2 se ha aislado de pacientes y es el producto final de las vacunas mRNA de Covid-19. En esta revisión se analiza cómo esta proteína puede disparar respuestas pro-inflamatorias y pro-coagulantes en cultivos primarios de dos tipos celulares muy afectados por SARS-Cov-2, como son los monocitos y las células endoteliales. La proteína S viral activa NF-κB, libera citoquinas pro-inflamatorias y activa el sistema del inflamasoma NLRP3, lo que produce la formación de IL-1β madura. Esto se acompaña de aumento de factores de coagulación, como el factor von Willebrand (vWF), el factor VIII o el factor tisular. Sin embargo, no incrementa los niveles de ADAMTS-13 para frenar la actividad pro-coagulante del vWF. Nuestros datos también indican que dichos efectos de la proteína S están mediador por receptores del tipo “toll-like”-4 en monocitos, pero no en el endotelio. Por otra parte, es conocido el papel del envejecimiento vascular prematuro y de la senescencia endotelial como factores de riesgo para la patología cardiovascular y aterotrombótica, complicaciones muy relevantes de la Covid-19 aguda o crónica. La proteína S del SARS-CoV-2 aumenta la senescencia celular y los marcadores de daño del DNA (β-galactosidasa asociada a senescencia, γH2AX), así como los efectores de parada celular (p53, p21, p16). También disminuye la disponibilidad de proteínas citoprotectoras, como la proteína anti-envejecimiento kloto, el factor Nrf2 y la hemoxigenasa-1. En conclusión, la proteína S actúa como estímulo pro-inflamatorio y pro-coagulantes en monocitos y endotelio humanos. Además, el tratamiento de las células con kloto recombinante o angiotensina-(1-7) protege de sus acciones pro-senescentes, pro-inflamatorias y pro-oxidantes. Globalmente, nuestros datos sugieren nuevos mecanismos patológicos para la Covid-19 y proponen dianas farmacológicas para prevenir sus complicaciones.
Abstract
Hyperinflammation, hypercoagulation, and endothelial injury are major findings in acute and post-Covid-19. The SARS-CoV-2 S protein has been detected as an isolated element in human tissues reservoirs and is the main product of mRNA Covid-19 vaccines. In this review, we analyzed whether the S protein alone triggers pro-inflammatory and pro-coagulant responses in primary cultures of two human cell types deeply affected by SARS-CoV-2, such are monocytes and endothelial cells. Indeed, viral S protein activated NF-κB, promoted pro-inflammatory cytokines release, and triggered the priming and activation of the NLRP3 inflammasome system resulting in mature IL-1β formation. This was paralleled by enhanced production of coagulation factors such as von Willebrand factor (vWF), factor VIII, or tissue factor. Additionally, S protein failed to enhance ADAMTS-13 levels to counteract the pro-coagulant activity of vWF multimers. Our data also indicate that these effects of the S protein are mediated by toll like-receptors-4 in monocytes, but not in endothelial cells. On the other hand, it is known that premature vascular aging and endothelial cell senescence are major risk factors for cardiovascular diseases and atherothrombotic disturbances, which are main complications of both acute and long Covid-19. The S protein of SARS-CoV2 enhanced cell senescence and DNA damage response markers (senescence-associated-β galactosidase, γH2AX), as well as growth-arrest effectors (p53, p21, p16). In parallel, the S protein reduced the availability of cytoprotective proteins, such as the anti-aging protein klotho, Nrf2 or heme oxygenase-1. In conclusion, S protein behaves both as a pro-inflammatory and pro-coagulant stimulus in human monocytes and endothelial cells. Furthermore, treating the cells with recombinant klotho or angiotensin-(1-7) protected against the S protein induced pro-senescence, pro-inflammatory, and pro-oxidant actions. Globally, our data proposes novel mechanisms of disease for Covid-19 and its vascular sequelae and provides pharmacological clues to prevent such complications.
Palabras clave: Proteína S del SARS-CoV-2; Células endoteliales; Monocitos; Inflamasoma NLRP3; Factores sanguíneos de coagulación; Senescencia celular.
Keywords: SARS-CoV-2 S Protein; Endothelial cells; Monocytes; NLRP3 inflammasome; blood coagulation Factors; Cell senescence.
INTRODUCCIÓN
La pandemia por el coronavirus 2019 (Covid-19) ha causado, desde su aparición en Wuhan, China, por encima de 750 millones de contagios y alrededor de siete millones de muertes en todo el mundo, según datos de la Organización Mundial de la Salud (1). El tremendo impacto de la pandemia y sus secuelas en la salud de la población y en las estructuras socioeconómicas mundiales, así como la necesidad de estar más preparados ante futuras pandemias, ha estimulado la investigación sobre los mecanismos fisiopatológicos de esta enfermedad.
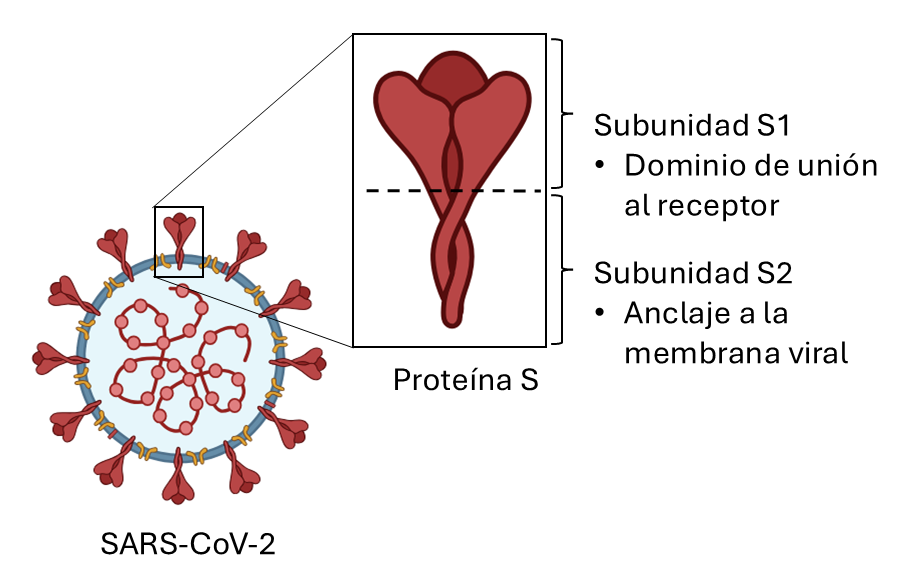
El agente causante (coronavirus 2 del síndrome respiratorio agudo severo, SARS-CoV-2) es un miembro de la familia de los coronavirus, recubierto por una “corona” de espículas (proteína S, del inglés “spike”), compuesta por dos dominios principales, S1 y S2 (Fig. 1) (2). La enzima convertidora de angiotensina 2 (ECA2) se identificó inicialmente como un receptor principal de la proteína S del virus, que permitía su entrada en las células (3). Sin embargo, otras moléculas de la superficie celular han sido relacionadas con el reconocimiento de la proteína S por distintos tipos de células huésped, incluyendo los receptores “toll-like” 4 (TLR4), basigina (CD147), o la dipeptidilpeptidasa-4 (DPP4/CD26) (4).
Trombosis y Covid-19
Desde el comienzo de la pandemia, la lesión del endotelio vascular en pacientes infectados por SARS-CoV-2 ha sido un hallazgo constante (5). Estudios histológicos postmortem han revelado inclusiones virales en células endoteliales apoptóticas, endotelitis linfocítica microvascular y la infiltración de células inmunes inflamatorias en la capa endotelial y en la zona perivascular, junto con la formación de microtrombos (6). Asimismo, una serie de observaciones clínicas ha identificado la pared vascular como uno de los principales tejidos afectados por el SARS-CoV-2 y responsable de las secuelas post Covid-19 (7).
Las respuestas inflamatorias en pacientes con Covid-19 severa se caracterizan por un reclutamiento intenso de células inmunes y elevados niveles de marcadores inflamatorios, incluyendo proteína C reactiva, ferritina y citoquinas, que se asocia a un estado de hiper-coagulación (8). En pacientes con Covid-19 se han encontrado niveles circulantes elevados de factores pro-coagulantes, como el factor von Willebrand (vWF) o el factor VIII (FVIII), producidos principalmente por las células endoteliales o el factor tisular (TF) secretado por células inmunes (9).
Sistema del inflamasoma NLRP3
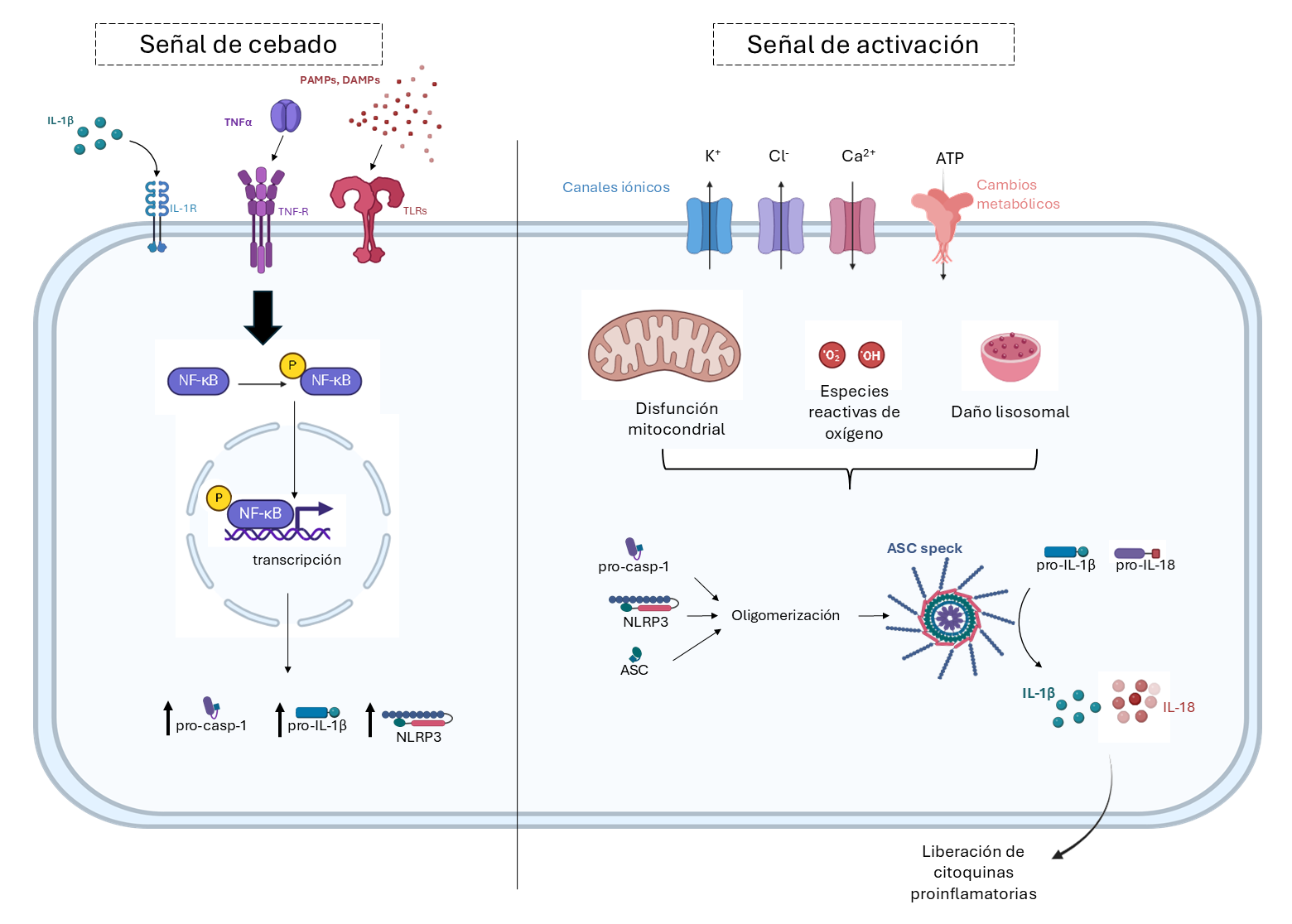
El sistema del inflamasoma NLRP3 (del inglés ”nucleotide-binding domain, leucine-rich repeat containing proteins”) es un sensor de primera línea en la respuesta inmune innata, considerado un factor clave en la inflamación vascular y la disfunción endotelial (10) con un papel relevante en múltiples patologías, como aterosclerosis y otras enfermedades cardiovasculares (11), diabetes mellitus (12) e infecciones virales como la Covid-1 (13). Tras una primera fase de producción de los componentes del inflamasoma, como NLRP3 o el precursor inactivo de la interleuquina (IL)-1β (pro-IL-1β), el inflamasoma requiere del ensamblaje de una estructura proteína multifuncional, que incluye NLRP3 y otras proteínas, como ASC (del inglés “adaptor molecule apoptosis-associated speck-like protein”). Posteriormente, este complejo activa la enzima caspasa-1, que escinde los precursores inactivos pro-IL-1β y pro-IL-18 en sus formas maduras activas (14) y, en monocitos, dispara la activación de gasdermina D, lo que favorece la formación de poros en la membrana celular y permiten la liberación de las citoquinas inflamatorias IL1-β e IL-18 (15) (Fig. 2). Un aspecto poco conocido en este sentido es el posible papel de la proteína S, detectada en suero y tejidos de pacientes tras la infección con SARS-CoV-2 (16) y que también es el producto final de las vacunas de mRNA frente al Covid-19.
síntesis
Efectos pro-inflamatorios de la proteína S del SARS-Cov-2 en cultivos de monocitos y endotelio humanos
Un primer hallazgo realizado por nuestro laboratorio has sido demostrar la capacidad de la proteína S del SARS-CoV-2 para estimular por sí misma vías pro-inflamatorias en cultivos primarios de monocitos y endotelio humanos. En estos dos tipos celulares, la proteína viral activa NF-κB, un regulador principal de la respuesta inflamatoria, y desencadena la señalización del inflamasoma NLRP3, además de inducir la síntesis de citoquinas soluble pro-inflamatorias y alterar el balance de reguladores de la coagulación (17).
La hiper-inflamación es una característica del Covid-19, y en sus complicaciones cardiovasculares, en la que los monocitos juegan un papel crucial, liberando citoquinas como IL-6, aumentada en pacientes con Covid-19, lo que se asocia a depleción de linfocitos T (18). En monocitos humanos, la proteína S también estimula, a través de la activación de NF-κB, la expresión de componentes del inflamasoma NLRP3 (19), facilitando el ensamblaje del complejo activo y la generación y liberación de IL-1β activa.
En monocitos y células endoteliales humanas, la IL-1β liberada es capaz de inducir una activación adicional del inflamasoma NLRP3 (17), de modo que al promover la síntesis y liberación de IL-1β, la proteína S inicia un bucle auto-inflamatorio que amplifica la producción local de esta citoquina en distintos tipos celulares.
Fisiopatológicamente, la sobre-activación del inflamasoma NLRP3 en las células vasculares se ha relacionado con enfermedades vasculares, como aterosclerosis, ictus o hipertensión y, más recientemente, con la vasculopatía e hiper-inflamación asociadas a la Covid-19 (20). Es decir, la proteína S del SARS-CoV-2 puede ser detectada por el sistema innato de defensa celular y producir IL-1β, una citoquina inflamatoria que se ha revelado como un eje crucial en la enfermedad vascular y la aterosclerosis. En este sentido, el estudio CANTOS demuestra que el bloqueo de la IL-1β con el anticuerpo monoclonal canakinumab reduce la inflamación crónica de bajo grado y la incidencia de eventos cardiovasculares en pacientes, independientemente de otros factores como la hiperlipidemia (21).
Por tanto, la producción local de IL-1β en células vasculares y monocitos humanos estimulados por la proteína S puede contribuir a la disfunción vascular en la Covid-19, favoreciendo la inflamación vascular y posiblemente amplificando y agravando lesiones vasculares previas.
Mecanismos pro-trombóticos de la proteína S del SARS-Cov-2 en cultivos de monocitos y endotelio humanos
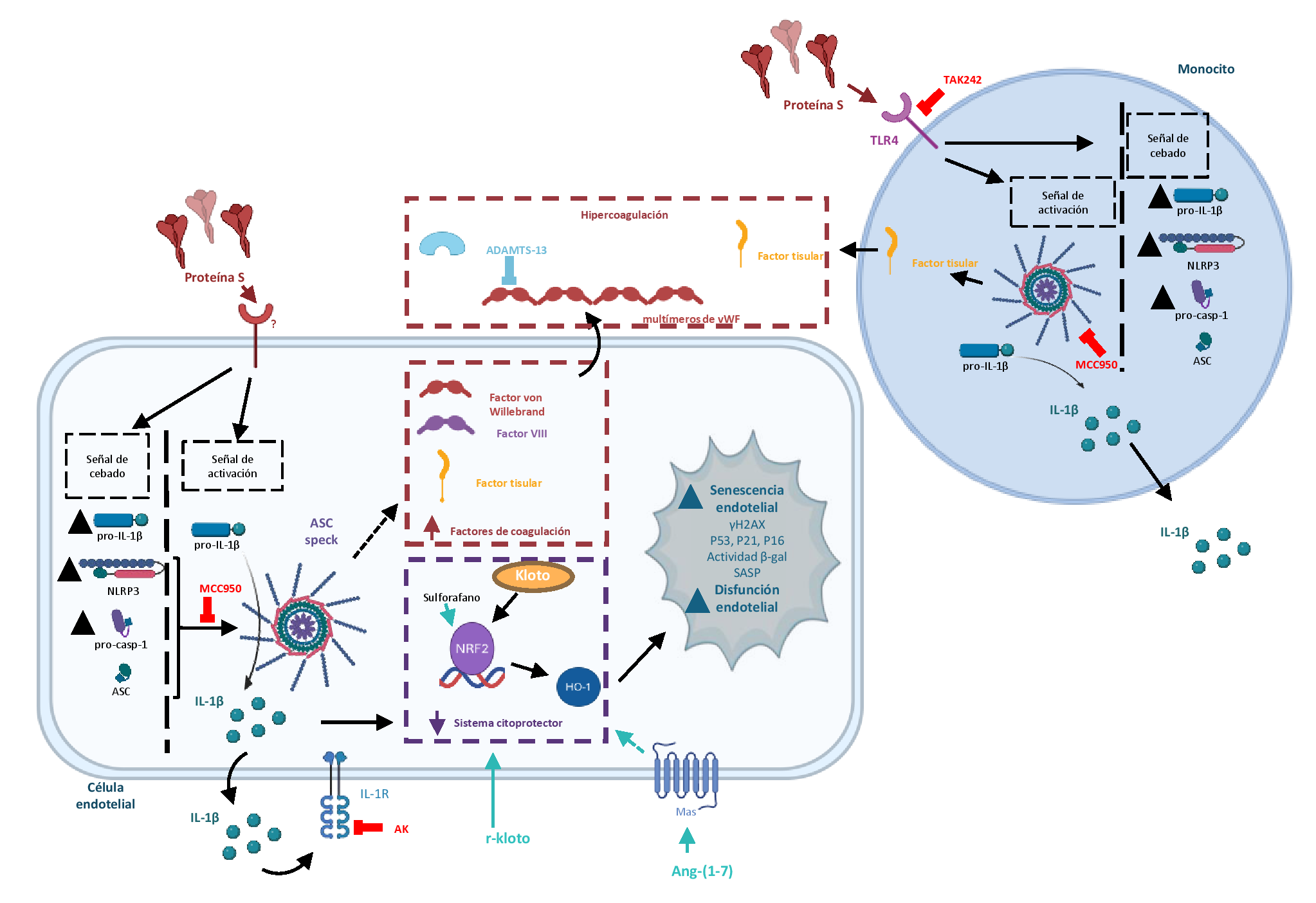
Estrechamente relacionados con la hiper-inflamación y la hipercoagulación, los eventos trombóticos son complicaciones importantes en pacientes con Covid-19 y sintomatología post-Covid-19 (22) y quizá también en los raros efectos adversos de las vacunas basadas en mRNA para la proteína S (23). De hecho, estas complicaciones post-vacuna se han relacionado con niveles circulantes elevados de proteína S (24). En este contexto, el factor de coagulación vWF, formado en células endoteliales y megacariocitos, está elevado en pacientes con Covid-19 (25), en los que constituye un marcador de endoteliopatía que predice un peor pronóstico (26). Tras su liberación, vWF se ensambla en multímeros filamentosos con alta actividad pro-coagulante y promueve la adhesión plaquetaria y la agregación (27). Para evitar la excesiva actividad trombogénica, la proteasa ADAMST-13, un regulador fisiológico de la hemostasia, recorta los multímeros de vWF en moléculas más pequeñas y menos activas (27). Nuestro laboratorio ha demostrado que, aunque tanto la proteína S como la IL-1β aumentan en contenido endotelial de vWF y su liberación al espacio extracelular, sólo la citoquina es capaz de inducir paralelamente la producción de ADAMST-13 que contrarresta la capacidad pro-coagulante de vWF (17). Es decir, en las células endoteliales humanas, la proteína viral S origina un disbalance en el ratio vWF:ADAMST-13 que favorece la formación de trombos, de modo similar a lo que se observa en algunas situaciones pro-coagulantes y pro-trombóticas, como la púrpura trombótica trombocitopénica o el ictus (27) . En este sentido, bajos niveles de ADAMST-13 se han descrito en pacientes con Covid-19, en los que un ratio vWF:ADAMST-13 elevado se correlaciona fuertemente con la severidad de la enfermedad, mientras que se asocia con endoteliopatía y disfunción inmune en pacientes con síndrome de Covid-19 persistente (28).
En paralelo, la proteína S incrementa también el contenido endotelial de otros factores implicados en las respuestas coagulantes, como FVIII y TF, igualmente asociados con la hiper-coagulabilidad en pacientes con Covid-19 (29). Estos factores se sobre-expresan asimismo en monocitos, donde la proteína S se comporta como las diferentes citoquinas pro-inflamatorias (IL-1β, IL-6, and TNF-α) liberadas en forma masiva en sujetos con Covid-19, lo que induce la expresión de TF en leucocitos y células no inmunológicas, favoreciendo un estado de hipercoagulación y la formación de trombos (29). La inhibición farmacológica del inflamasoma NLRP3 o el bloqueo de los IL-1R tienden a atenuar la sobre-expresión endotelial de factores de coagulación inducidos por la proteína S (27). Por su parte, en monocitos, el inflamasoma NLRP3 media la liberación de TF, que es un iniciador primario de la cascada de coagulación (30). Así pues, parece existir una íntima relación entre las actividades pro-inflamatorias y pro-coagulantes de la proteína S del SARS-Cov-2, lo que permite proponer diferentes intervenciones farmacológicas para interferir con los efectos deletéreos de esta partícula viral en células inmunes y vasculares humanas (Fig. 1). Apoyando esta idea, se ha descrito en monocitos de pacientes con Covid-19 severa una sobre-activación espontánea del inflamasoma NLRP3, con secreción de IL-1β que puede ser revertida mediante el tratamiento de los pacientes con el bloqueante de los receptores para IL-1 anakinra (31).
Posibles receptores implicados en los efectos de la proteína S del SARS-CoV-2
Aunque la ECA2 se ha identificado como un receptor relevante para la espícula del SARS-Cov-2 en muchos tipos celulares (3), no se ha detectado en cultivos primarios de monocitos humanos, ni en nuestro laboratorio (17), ni en otros estudios publicados (32). Tampoco hemos observado este receptor en células endoteliales humanas (27), al igual que otros investigadores (33). Otros receptores que pueden favorecer el reconocimiento y la interacción de la proteina S con las células huésped incluyen los receptores “toll-like” tipo 4 (TLR4), basigina (CD147) y dipeptidilpeptidasa-4 (DDP4/CD26) (4, 32). Los monocitos expresan constitutivamente receptores TLR4 de superficie, que es el receptor canónico implicado en el reconocimiento de lipopolisacáridos bacterianos y ha sido asociado con diversas patologías (34). La unión del ligando al receptor TLR4 lleva a su oligomerización que activa el factor 88 de diferenciación mieloide (MYD88) y conlleva la subsecuente activación del factor de transcripción NF-κB (35). Por eso, el bloqueo de TLR4 ha sido propuesto como un posible tratamiento para pacientes de Covid-19 (36), lo que puede ser una opción interesante para atenuar las acciones directas de la proteína s como elemento viral aislado.
Además de los episodios agudos de Covid-19, la proteína S puede tener un papel en las secuelas crónicas el SARS-CoV-2, que se han relacionado con la persistencia de niveles circulantes elevados de la misma (37). Nuestra hipótesis es que una inflamación sostenida del sistema inmune y del endotelio, junto a la hipercoagulación y la trombosis mediadas por la proteína S contribuye a las secuelas crónicas de la enfermedad. Dado que los vasos sanguíneos llegan a todos los órganos y las células inmunes están presentes en todos los tejidos, ambos tipos celulares pueden ser el mecanismo común de la amplia variedad de síntomas del Covid-19 persistente. Asimismo, en su conjunto, nuestros datos apoyan el papel etiopatogénico de la proteína S en las manifestaciones clínicas del Covid-19, tango agudas como crónicas, y sugieren que las intervenciones farmacológicas basadas en el bloqueo de esta proteína, en la inhibición del inflamasoma NLRP3, en los anticuerpos monoclonales o proteínas de fusión frente a IL-1β o IL-1R, así como los antagonistas de TLR4, pueden ser alternativas terapéuticas válidas, junto a los tratamientos anticoagulantes.
Efecto pro-senescente de la proteína S del SARS.Cov-2 en endotelio humano
Otro hallazgo importante realizado en nuestro laboratorio es la observación de que la proteína S es capaz de inducir marcadas respuestas pro-senescentes en cultivos de células endoteliales humanas (38). Conviene recordar que el envejecimiento vascular prematuro es un aspecto muy ligado al desarrollo de patología cardiovascular y suele ser un buen marcador de fragilidad individual (39). La senescencia celular es uno de los aspectos implicados en el envejecimiento vascular y que favorecen el daño tisular. Las células endoteliales senescentes sufren una parada irreversible del ciclo celular y adquieren un fenotipo senescente, secretor, pro-oxidante y pro-inflamatorio (SASP), que comparte características con el endotelio disfuncional (40). Las células endoteliales con este fenotipo SASP liberan citoquinas, como IL-6 o IL-1β, y otros factores oxidantes, inflamatorios y trombóticos que modifican el estado redox celular y propagan la senescencia a las células vecinas, favoreciendo la progresión de la enfermedad vascular (41). Además, las células endoteliales senescentes sobre-expresan el factor vWF y favorecen un ambiente trombogénico (42), análogo al observado en Covid-19 aguda y persistente (43).
El envejecimiento, uno de los factores de riesgo más importantes en los pacientes de Covid-19, se asocia en general a un declive de las respuestas de defensa intracelular, mientras la senescencia endotelial se caracteriza por una reducción en la capacidad antioxidante y en la homeostasis redox (44). Entre los principales factores que confieren protección a las células, el factor eritroide 2 (Nrf2) es un sistema citoprotector conservado evolutivamente que se considera un potente modulador de la longevidad en distintas especies y un mecanismo molecular que relaciona el estrés oxidativo con el envejecimiento (45). Nfr2 protege frente a estímulos oxidantes promoviendo la expresión de genes que codifican proteínas anti-oxidantes y anti-inflamatorias, como la hemoxigenasa-1 (HO-1). Esta molécula proporciona protección celular degradando grupos hemo pro-oxidantes para formar bilirrubina y monóxido de carbono como señalizador (45).
En nuestro laboratorio, hemos descrito recientemente la capacidad que tiene la proteína S del SARS-Cov-2 para debilitar este sistema protector en células endoteliales humanas (38). Una posible explicación para este efecto deletéreo puede ser la disminución de los niveles celulares de la proteína kloto producida por la presencia de la proteína S viral (38). En células endoteliales, kloto induce la expresión de Nfr2, protegiéndolas de agresiones derivadas de estrés oxidativo, mientras que tiene efectos anti-apoptóticos y anti-senescentes (46). Secundando esta hipótesis, la administración de kloto recombinante (r-kloto) en cultivos de endotelio humano revierte las acciones pro-senescentes de la proteina S (38). Asimismo, el tratamiento con angiotensina-(Ang)-(1-7) produce efectos similares, lo que se explica porque este péptido, componente del brazo protector de sistema renina-angiotensina, aumenta los niveles intracelulares de kloto y, consecuentemente, de Nrf2 y HO-1 en células endoteliales (38, 46). Además, tanto kloto como Ang-(1-7) favorecen la secreción de óxido nítrico endotelial (47, 48) y corrigen las alteraciones de las repuestas vasodilatadoras dependientes de endotelio producidas por la proteína S (38). La capacidad de estos compuestos para prevenir la senescencia endotelial originada por diferentes estímulos ya había sido previamente demostrada por nuestro grupo (46), lo que sugiere un posible papel como agentes senoterapéuticos. En este sentido, más que eliminar las células senescentes, estos fármacos se comportan como senostáticos, capaces de prevenir el desarrollo de senescencia endotelial y la consecuente secreción de componentes pro-inflamatorios asociados al SASP, incluyendo los originados por la proteína S del SARS-Cov-2.
CONCLUSIONES
De los resultados expuestos, podemos concluir que la proteína S del SARS-CoV-2 estimula directamente células endoteliales e inmunes de origen humano, a través de receptores no claramente determinados, promoviendo la activación de mecanismos pro-inflamatorios, pro-coagulantes y pro-senescentes. Ello nos permite emitir la hipótesis de que la interferencia de la proteína S con su receptor eventual o con sus respectivos mecanismos de señalización puede tener implicaciones terapéuticas en las complicaciones agudas o crónicas del Covid-19, así como en los efectos adversos derivados de vacunas mRNA que expresan esta proteína.
Además, hemos demostrado la capacidad de la proteína S para activar el inflamasoma NLRP3, lo que está relacionado con la sobre-expresión de factores pro-coagulantes, así como con el agotamiento de los sistemas de defensa citoprotectora frente a la senescencia endotelial prematura inducida. Globalmente considerada, la activación del inflamasoma NLRP3 emerge como un mecanismo central en el efecto deletéreo de la proteína S del SARS-CoV-2 sobre el endotelio. La citoquina recombinante IL-1β mimetiza las acciones de la proteina S, lo que tiene relevancia fisiopatológica y terapéutica, ya que esta citoquina, producto principal del inflamasoma NLRP3, se ha relacionado con la patología vascular humana en el estudio CANTOS. De hecho, el SARS-CoV-2 activa la vía IL-1/IL-6 en mayor medida que otros coronavirus (49). Además, productos derivados del inflamasoma NLRP3, como IL-1β e IL-18 están incrementados en pacientes con Covid-19 aguda severa y correlacionan positivamente con consecuencias clínicas adversas (50).
Finalmente, nuestros resultados indican que ciertos elementos aislados persistentes del SARS-CoV-2, como la proteína S de la corona viral, produce daños pró-inflamatorios y pro-oxidantes en los vasos y promueve senescencia prematura y disfunción vascular (Fig. 3). Como aspectos terapéuticos a tener en consideración, cabe destacar el bloqueo de la sobre-activación del inflamasoma NLRP3 y la excesiva generación de sus productos finales, así como el empleo de fármacos como kloto y Ang-(1-7), que pueden restaurar la capacidad citoprotectora vascular, y pueden constituir alternativas farmacológicas valiosas para el tratamiento de las secuelas derivadas del Covid-19.
AGRADECIMIENTOS
Este trabajo ha sido financiado por la Unión Europea (REACT-EU-Comunidad de Madrid y European Regional Development Fund; SPACE2-CV-COVID-CM), el Plan Nacional de I+D (PID2020-115590RB-100/AEI/https://doi.org/10.13039/ 501100011033; PID2021-126274OB-I00; PID2022-137373OB-I00-MICIU/AEI/ 10.13039 /501100011033/FEDER, UE y PID2023-147378OB-I00-MCIN/AEI/10.13039/501100011033/ FEDER, UE), y el Fondo de Investigación Sanitaria-FIS Carlos III (PI20/00923). L.S. tiene un contrato FPI Universidad Autónoma de Madrid (SFPI / 2020-00053). IV tiene un contrato Sara Borrell postdoctoral (CD22/00101). AV tiene un contrato Sara Borrell postdoctoral (CD24/00217)
DECLARACIÓN DE TRANSPARENCIA
Los autores/as de este artículo declaran no tener ningún tipo de conflicto de intereses respecto a lo expuesto en el presente trabajo.
BIBLIOGRAFÍA
- ↑World Health Organization. COVID-19 epidemiological update (Edition 160). 2023. Available from: https:// www. who. int/ publi catio ns/m/ item/ covid‑ 19‑ epidemiological‑update‑‑‑ 27‑ october‑ 2023.
- ↑Watanabe Y, Allen JD, Wrapp D, et al. Site-specific glycan analysis of the SARS-CoV-2 spike. Science. 2020;369:330–333. doi: 10.1126/science.abb9983
- ↑Lan J, Ge J, Yu J, et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature. 2020;581:215–220. doi: 10.1038/s41586-020-2180-5
- ↑Valencia I, Peiro C, Lorenzo O, et al. DPP4 and ACE2 in diabetes and COVID-19: therapeutic targets for cardiovascular complications? Front Pharmacol. 2020;11:1161. doi: 10.3389/fphar.2020.01161
- ↑Gu SX, Tyagi T, Jain K, et al. Thrombocytopathy and endotheliopathy: crucial contributors to COVID-19 thromboinflammation. Nat Rev Cardiol. 2021;18:194–209. doi: 10.1038/s41569-020-00469-1
- ↑Varga Z, Flammer AJ, Steiger P, et al. Endothelial cell infection and endothelitis in COVID-19. Lancet. 2020;395:1417–1418. doi: 10.1016/S0140-6736(20)30937-5
- ↑Martinez-Salazar B, Holwerda M, Studle C, et al. COVID-19 and the vasculature: current aspects and long-term consequences. Front Cell Dev Biol.2022;10:824851. doi: 10.3389/fcell.2022.824851
- ↑Moore JB, June CH. Cytokine release syndrome in severe COVID-19. Science. 2020;368:473–474. doi: 10.1126/science.abb8925
- ↑Iba T, Levy JH, Connors JM, et al. The unique characteristics of COVID-19 coagulopathy. Crit Care. 2020;24:360. doi: 10.1186/s13054-020-03077-0
- ↑Bai B, Yang Y, Wang Q, et al. NLRP3 inflammasome in endothelial dysfunction. Cell Death Dis.2020;11:776. doi: 10.1038/s41419-020-02985-x
- ↑Toldo S, Mezzaroma E, Buckley LF, et al. Targeting the NLRP3 inflammasome in cardiovascular diseases. Pharmacol Ther. 2022;236:108053. doi: 10.1016/j.pharmthera.2021.108053
- ↑Ding S, Xu S, Ma Y, et al. Modulatory mechanisms of the NLRP3 Inflammasomes in diabetes. Biomolecules. 2019;9:850. doi: 10.3390/biom9120850
- ↑Zhao C, Zhao W. NLRP3 Inflammasome-a key player in antiviral responses. Front Immunol. 2020;11:211. doi: 10.3389/fimmu.2020.00211
- ↑He Y, Hara H, Nuñez G. Mechanism and regulation of NLRP3 Inflammasome activation. Trends Biochem Sci. 2016;41: 1012–1021. doi: 10.1016/j.tibs.2016.09.002
- ↑Lamkanfi M, Dixit VM. Mechanisms and functions of inflammasomes. Cell. 2014;157:1013–1022. doi: 10.1016/j.cell.2014.04.007
- ↑Perico L, Morigi M, Galbusera M, et al. SARS-CoV-2 spike protein 1 activates microvascular endothelial cells and complement system leading to platelet aggregation. Front Immunol. 2022;13: 827146. doi: 10.3389/fimmu.2022.827146
- ↑Villacampa A, Alfaro E, Morales C, et al. SARS-CoV-2 S protein activates NLRP3 inflammasome and deregulates coagulation factors in endothelial and immune cells. Cell Commun Signal. 2024 Jan 15;22(1):38. doi: 10.1186/s12964-023-01397-6.
- ↑Merad M, Martin JC. Pathological inflammation in patients with COVID-19: a key role for monocytes and macrophages. Nat Rev Immunol. 2020; 20:355–362. doi: 10.1038/s41577-020-0331-4
- ↑Bauernfeind FG, Horvath G, Stutz A, et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J Immunol. 2009;183: 787–791. doi: 10.4049/jimmunol.0901363
- ↑van den Berg DF, Te Velde AA. Severe COVID-19: NLRP3 Inflammasome dysregulated. Front Immunol. 2020; 11:1580. doi: 10.3389/fimmu.2020.01580
- ↑Ridker PM, Everett BM, Thuren T, et al. Antiinflammatory therapy with canakinumab for atherosclerotic disease. N Engl J Med. 2017; 377:1119–1131. doi: 10.1056/NEJMoa1707914
- ↑Jose RJ, Manuel A. COVID-19 cytokine storm: the interplay between inflammation and coagulation. Lancet Respir Med. 2020; 8:e46–7. doi: 10.1016/S2213-2600(20)30216-2. Epub 2020 Apr 27.
- ↑Yasmin F, Najeeb H, Naeem U, et al. Adverse events following COVID-19 mRNA vaccines: a systematic review of cardiovascular complication, thrombosis, and thrombocytopenia. Immun Inflamm Dis. 2023; 11:e807. doi: 10.1002/iid3.807
- ↑Yonker LM, Swank Z, Bartsch YC, et al. Circulating spike protein detected in post-COVID-19 mRNA vaccine myocarditis. Circulation. 2023; 147:867–876. doi: 10.1161/CIRCULATIONAHA.122.061025
- ↑Ward SE, Curley GF, Lavin M, et al. Von Willebrand factor propeptide in severe coronavirus disease 2019 (COVID-19): evidence of acute and sustained endothelial cell activation. Br J Haematol. 2021; 192:714–719. doi: 10.1111/bjh.17273
- ↑Wibowo A, Pranata R, Lim MA, et al. Endotheliopathy marked by high von Willebrand factor (vWF) antigen in COVID-19 is associated with poor outcome: a systematic review and meta-analysis. Int J Infect Dis. 2022; 117:267–273. doi: 10.1016/j.ijid.2021.06.051
- ↑Santamaria S, de Groot R. ADAMTS proteases in cardiovascular physiology and disease. Open Biol. 2020; 10:200333. doi: 10.1098/rsob.200333
- ↑Marco A, Marco P. Von Willebrand factor and ADAMTS13 activity as clinical severity markers in patients with COVID-19. J Thromb Thrombol. 2021; 52:497–503. doi: 10.1007/s11239-021-02457-9
- ↑Sachetto TAA, Mackman N. Tissue factor and COVID-19: an update. Curr Drug Targets. 2022; 23:1573–1577. doi: 10.2174/1389450123666220926144432
- ↑Zelaya H, Rothmeier AS, Ruf W. Tissue factor at the crossroad of coagulation and cell signaling. J Thromb Haemost. 2018; 16:1941–1952. doi: 10.1111/jth.14246
- ↑Bertoni A, Penco F, Mollica H, et al. Spontaneous NLRP3 inflammasome-driven IL-1β secretion is induced in severe COVID-19 patients and responds to anakinra treatment. J Allergy Clin Immunol. 2022; 150:796–805. doi: 10.1016/j.jaci.2022.05.029
- ↑Radzikowska U, Ding M, Tan G, et al. Distribution of ACE2, CD147, CD26, and other SARS-CoV-2 associated molecules in tissues and immune cells in health and in asthma, COPD, obesity, hypertension, and COVID-19 risk factors. Allergy. 2020; 75:2829–2845. doi: 10.1111/all.14429
- ↑McCracken IR, Saginc G, He L, et al. Lack of evidence of angiotensin-converting enzyme 2 expression and replicative infection by SARS-CoV-2 in human endothelial cells. Circulation. 2021; 143:865–868. doi: 10.1161/CIRCULATIONAHA.120.052824
- ↑den Dekker WK, Cheng C, et al. Toll like receptor 4 in atherosclerosis and plaque destabilization. Atherosclerosis. 2010; 209: 314–320. doi: 10.1016/j.atherosclerosis.2009.09.075
- ↑Lu YC, Yeh WC, Ohashi PS. LPS/TLR4 signal transduction pathway. Cytokine. 2008; 42:145–151. doi: 10.1016/j.cyto.2008.01.006
- ↑Kaushik D, Bhandari R, Kuhad A. TLR4 as a therapeutic target for respiratory and neurological complications of SARS-CoV-2. Expert Opin Ther Targets. 2021; 25:491–508. doi: 10.1080/14728222.2021.1918103
- ↑Swank Z, Senussi Y, Manickas-Hill Z, et al. Persistent circulating severe acute respiratory syndrome coronavirus 2 spike is associated with post-acute coronavirus disease 2019 sequelae. Clin Infect Dis. 2023; 76: e487–90. doi: 10.1093/cid/ciac722
- ↑Villacampa A, Shamoon L, Valencia I, et al. SARS-CoV-2 S protein reduces cytoprotective defenses and promotes human endothelial cell senescence. Aging Dis. 2024 Jul 5. doi: 10.14336/AD.2024.0405.
- ↑Saz-Lara A, Cavero-Redondo I, Pascual-Morena C, et al. Early vascular aging as an index of cardiovascular risk in healthy adults: confirmatory factor analysis from the EVasCu study. Cardiovasc Diabetol. 2023 Aug 17; 22(1):209. doi: 10.1186/s12933-023-01947-9.
- ↑Khavinson V, Linkova N, Dyatlova A, et al. Senescence-associated secretory phenotype of cardiovascular system cells and inflammaging: perspectives of peptide regulation. Cells. 2022 Dec 27;12(1):106. doi: 10.3390/cells12010106.
- ↑Birch J, Gil J. Senescence and the SASP: Many therapeutic avenues. Genes Dev. 2020 Dec 1; 34(23-24):1565-1576. doi: 10.1101/gad.343129.120
- ↑Bochenek ML, Schütz E, Schäfer K. Endothelial cell senescence and thrombosis: Ageing clots. Thromb Res. 2016: 147:36-45. doi: 10.1016/j.thromres.2016.09.019.
- ↑Valencia I, Lumpuy-Castillo J, Magalhaes G, et al. Mechanisms of endothelial activation, hypercoagulation and thrombosis in COVID-19: a link with diabetes mellitus. Cardiovasc Diabetol. 2024; 23(1):75. doi: 10.1186/s12933-023-02097-8.
- ↑Han Y, Kim SY. Endothelial senescence in vascular diseases: current understanding and future opportunities in senotherapeutics. Exp Mol Med. 2023;55(1): 1-12. doi: 10.1038/s12276-022-00906-w.
- ↑Yu C, Xiao JH. The Keap1-Nrf2 System: A mediator between oxidative stress and aging. Oxid Med Cell Longev. 2021:2021: 6635460. doi: 10.1155/2021/6635460.
- ↑Romero A, San Hipólito-Luengo Á, Villalobos LA, et al. The angiotensin-(1-7)/Mas receptor axis protects from endothelial cell senescence via klotho and Nrf2 activation. Aging Cell. 2019;18(3):e12913. doi: 10.1111/acel.12913.
- ↑Peiró C, Vallejo S, Gembardt F, et al. Complete blockade of the vasorelaxant effects of angiotensin-(1-7) and bradykinin in murine microvessels by antagonists of the receptor Mas. J Physiol. 2013;591(9): 2275-85. doi: 10.1113/jphysiol.2013.251413.
- ↑Richter B, Haller J, Haffner D, et al. Klotho modulates FGF23-mediated NO synthesis and oxidative stress in human coronary artery endothelial cells. Pflugers Arch. 2016;468(9): 1621-35. doi: 10.1007/s00424-016-1858-x.
- ↑Blanco-Melo D, Nilsson-Payant BE, Liu WC, et al. Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell. 2020;181(5):1036-1045.e9. doi: 10.1016/j.cell.2020.04.026.
- ↑Rodrigues TS, de Sá KSG, Ishimoto AY, et al. Inflammasomes are activated in response to SARS-cov-2 infection and are associated with COVID-19 severity in patients. J Exp Med. 2021;218(3):e20201707. doi: 10.1084/jem.20201707.
ranm tv
Carlos F. Sánchez Ferrer
Universidad Autónoma de Madrid
C/ Arzobispo Morcillo 4 · 28029 Madrid
Tlf.: +34 636 348 399 | E-Mail: carlosf.sanchezferrer@uam.es
An RANM. 2025;142(01): 73-81
Enviado*: 11.06.24
Revisado: 17.06.24
Aceptado: 19.07.24
* Fecha de lectura en la RANM