

Resumen
El objetivo principal de nuestros estudios es comprender las bases neuronales subyacentes a las dificultades fonológicas en la dislexia. En primer lugar, revisaremos el marco teórico de investigación generado en torno a la teoría fonológica de la dislexia. En segundo lugar, repasaremos cuales son los mecanismos neuronales involucrados en la segmentación del habla en lectores controles. En este apartado demostraremos que la sincronización entre los ritmos del habla y las oscilaciones neuronales a distintas bandas de frecuencia juega un papel clave en la segmentación del habla. A continuación, presentaremos distintos estudios que sugieren que los lectores disléxicos, presentan una desincronización entre los ritmos del habla y las oscilaciones neuronales en regiones auditivas. Esta falta de sincronización podría causar los problemas de percepción auditiva y las dificultades fonológicas que observamos en los lectores con dislexia. Finalmente, presentaremos estudios recientes de nuestro laboratorio que apoyan la teoría de la desincronización neuronal en la dislexia y muestran que estos problemas también están presentes en los niños con dislexia.
Abstract
The main objective of our studies is to understand the neural bases underlying phonological difficulties in dyslexia. First, we will review the theoretical research framework generated around the phonological theory of dyslexia. Second, we will review what are the neural mechanisms involved in the segmentation of speech in control readers. In this section we will demonstrate that the synchronization between speech rhythms and neural oscillations at different frequency bands plays a key role in the segmentation of speech. Next, we will present different studies that suggest that dyslexic readers present a desynchronization between speech rhythms and neuronal oscillations in auditory regions. This lack of synchronization could cause the auditory perception problems and the phonological difficulties that we observe in readers with dyslexia. Finally, we will present recent studies from our laboratory that support the theory of neuronal desynchronization in dyslexia and show that these problems are also present in children with dyslexia.
Palabras clave: Dislexia; Trastorno fonológico; Ritmos del habla; Oscilaciones neuronales; Desincronización neuronal; Magnetoencefalografía.
Keywords: Dyslexia; Phonological disorder; Speech rhythms; Neural oscillations; Neuronal desynchronization; Magnetoencephalography.
La teoría fonológica de la dislexia
La dislexia es un trastorno neurológico con una base genética que afecta a la adquisición y el procesamiento del lenguaje escrito. La dislexia se manifiesta en la dificultad para aprender a leer a pesar de tener una inteligencia adecuada, no mostrar deficiencias sensoriales y haber disfrutado de oportunidades socioculturales. Afecta aproximadamente al 5-7% de la población y parece ser más frecuente entre los hombres que entre las mujeres (Finucci & Childs, 1981). Las causas de la dislexia no son conocidas, aunque existen distintas teorías al respecto (Ramus et al., 2003). Actualmente, la explicación predominante a nivel cognitivo sobre la causa de la dislexia es la teoría fonológica (Ramus et al., 2003). Este enfoque postula que los lectores disléxicos tienen un deterioro específico en la representación, acceso y/o recuperación de los sonidos del habla. Los déficits fonológicos en la dislexia se reflejan clásicamente en: (1) la falta de conciencia fonológica, esto es, dificultades en la capacidad de identificar y manipular los sonidos del lenguaje (prosodia, sílabas y fonemas); (2) una pobre memoria verbal a corto plazo, esto es, una habilidad débil para mantener momentáneamente activa las representaciones fonológicas; (3) la recuperación léxica lenta, esto es, un retardo en la habilidad de recuperar las formas de las palabras de la memoria a largo plazo (Wagner & Torgesen, 1987; Ramus, 2004; Vellutino et al., 2004).
Actualmente, el interés de los investigadores se centra en identificar el origen del déficit fonológico presente en los lectores disléxicos.
Sincronización entre los ritmos del habla y ritmos cerebrales
Durante el discurso, las modulaciones temporales del habla fluctúan a distintos ritmos (Figura 1) (Arvaniti, 2009; Ghitza & Greenberg, 2009). Los ritmos más lentos, corresponden a la información prosódica que cambia cada 0.5 − 2 segundos. Las palabras y las sílabas aparecen en el flujo del habla a tasas relativamente constantes, cada 4 − 7 veces por segundo. Por último, los fonemas se producen a un ritmo de entre 25 y 40 veces por segundo. La regularidad temporal de las sucesivas unidades fonológicas modula de forma casi rítmica la envolvente del habla. El ritmo característico de cada unidad lingüística varía para cada idioma, pero siempre dentro de los límites indicados anteriormente.
El ritmo de un idioma, además de captar y mantener la atención del oyente, proporciona pistas para la segmentación y comprensión del mensaje a transmitir. La segmentación del habla es el proceso de identificar los límites entre palabras, sílabas o fonemas en el habla. Estudio previos en adultos han demostrado que la segmentación del habla se realiza a través de la sincronización entre las oscilaciones neuronales en regiones auditivas y los distintos ritmos del habla (Figura 1) (Poeppel, 2003; Hickok & Poeppel, 2007; Ghitza & Greenberg, 2009; Giraud & Poeppel, 2012; Bourguignon et al., 2013; Gross et al., 2013; Molinaro & Lizarazu, 2018). Las oscilaciones neuronales en la banda frecuencial delta (0.5 – 2 Hz) se sincronizan con la información sobre la estructura de frases y la prosodia. Las oscilaciones neuronales en la banda de frecuencia theta (4 – 7 Hz) se sincronizan con las modulaciones temporales asociadas a las palabras y sílabas. Finalmente, las oscilaciones neuronales en la banda de frecuencia gamma (25 – 40 Hz) se sincronizan con las modulaciones rápidas del habla asociadas a la información de los fonemas. Es importante destacar que las regiones auditivas del hemisferios izquierdo y derecho desempeñan diferentes roles en la segmentación del habla: mientras que las regiones auditivas del hemisferio derecho están especializadas en el procesamiento de modulaciones delta y theta, las regiones auditivas del hemisferio izquierdo están asociadas con el procesamiento de fluctuaciones gamma (Poeppel, 2003). El procesamiento en paralelo del habla a diferentes escalas temporales permite que las representaciones sensoriales de los sonidos sean estables a pesar de la presencia de ruido (Greenberg & Arai, 2001; Saberi & Perrot, 1999) y aumenta la capacidad de segmentación de las respuestas neuronales (Panzeri et al., 2010).
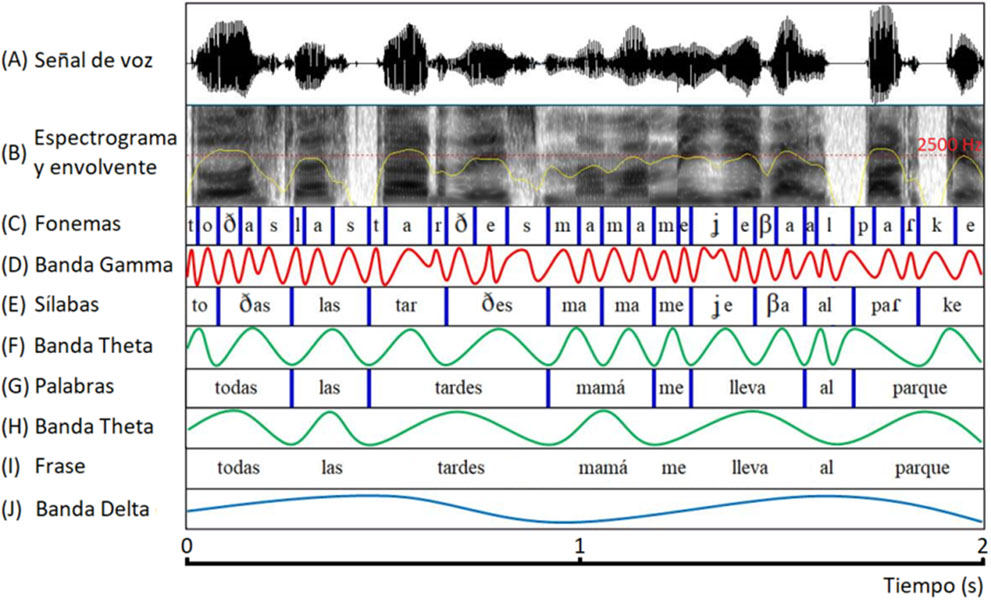
Sincronización entre los ritmos del habla y ritmos cerebrales en la dislexia
Numerosos estudios sugieren que los trastornos fonológicos en la dislexia son el resultado de dificultades en el procesamiento perceptivo de los sonidos (Tallal, 1980; De Martino et al., 2001; Amitay, Ahissar & Nelken, 2002; Goswami et al., 2002; Rey et al., 2002). En general, es razonable suponer que una mala percepción auditiva puede afectar a la segmentación del habla y conducir a representaciones fonológicas menos precisas en la dislexia (Goswami, 2011; Giraud & Poeppel, 2012). Estudios recientes indican que la desincronización entre los ritmos del habla y las oscilaciones neuronales puede estar relacionada con las dificultades en la segmentación del habla y los problemas fonológicos presentes en la dislexia (Abrams et al., 2009; Goswami, 2011; Giraud & Poeppel, 2012). La Tabla 1 resume los estudios de neuroimagen existentes, indicando de la manera más precisa posible los resultados en función de la técnica de neuroimagen, el tipo de estímulo, el tipo de medición y el método de análisis.
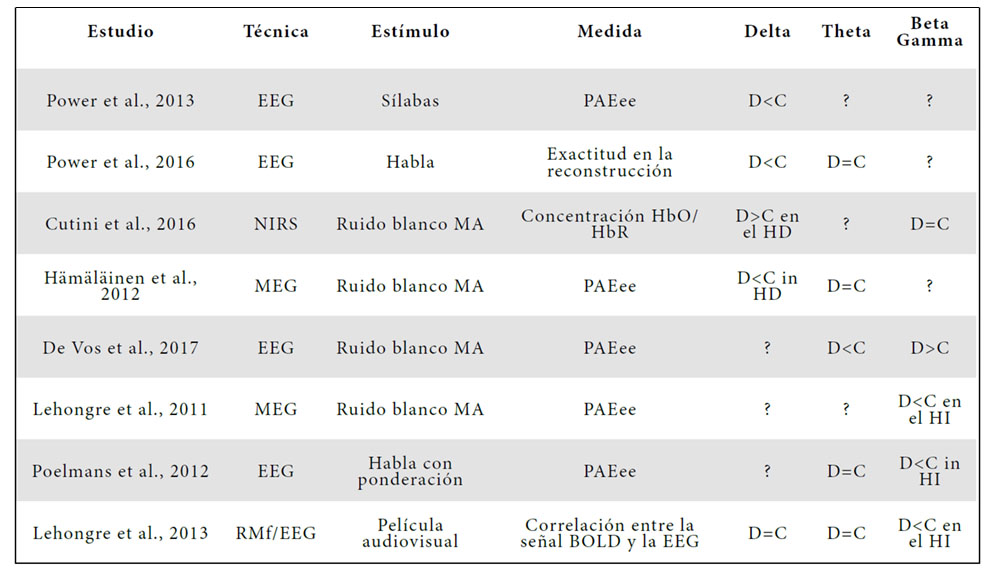
Por un lado, Goswami (2011) ha planteado la hipótesis de que los lectores disléxicos muestran una desincronización neuronal en las bandas frecuenciales delta y theta, lo que conduce a déficits en la codificación de la información a nivel de la prosodia, las palabras y las sílabas. Power y sus colaboradores (2013, 2016) mostraron que el cerebro disléxico muestra una sincronización más débil en la banda delta durante el procesamiento del habla en las regiones auditivas del hemisferio derecho. Esta falta de sincronización en la banda delta también se ha observado durante el procesamiento de señales auditivas no lingüísticas en participantes con dislexia (Hämäläinen et al., 2012). Hämäläinen y sus colaboradores (2012), midieron los potenciales evocados auditivos de estado estable (PAEee) durante la presentación de ruido blanco modulado en amplitud a 2, 4, 10 y 20 Hz en lectores con y sin dislexia. Los resultados de este estudio mostraron que los lectores disléxicos exhibían una respuesta PAEee más débil a los 2 Hz (ritmos prosódicos) en las regiones auditivas del hemisferio derecho. Más recientemente, utilizando la espectroscopia de infrarrojo cercano o NIRS (near-infrared spectroscopy), Cutini y sus colaboradores (2016) revelaron que lectores disléxicos mostraban índices atípicos de concentración de HbO (hemoglobina oxigenada) durante el procesamiento de ruido blanco modulado a 2 Hz en el giro supramarginal derecho, una de las región involucradas en el procesamiento del ritmo y la prosodia del habla (Geiser et al., 2008). En la banda theta, la evidencia de una desincronización neuronal en lectores con dislexia no es concluyente. De Vos y sus colaboradores (2017), mostraron una disminución de la respuesta PAEee en lectores durante el procesamiento de ruido blanco modulado a 4 Hz (ritmo de palabras y sílabas) en regiones auditivas bilaterales. Sin embargo, no se han encontrado indicios de una falta de sincronización entre las oscilaciones neuronales en la banda theta y los ritmos silábicos en el habla (Lehongre et al., 2013; Power et al., 2016).
Por otro lado, Giraud y Poeppel (2012) han planteado la hipótesis de que los lectores disléxicos podrían mostrar una desincronización neuronal en la banda gamma, lo que podría afectar el procesamiento de las unidades fonémicas. Lehongre y sus colaboradores (2011) midieron la respuesta PAEee a ruido blanco modulado en amplitud en un rango de 10 a 80 Hz y mostraron que la respuesta PAEee a 30 Hz (ritmos fonémicos) era más débil en las regiones auditivas del hemisferio izquierdo para los lectores disléxicos en comparación con los controles. De manera similar, utilizando la electroencefalografía (EEG), Poelmans y sus colaboradores (2012) encontraron un decrecimiento de la respuesta PAEE durante el procesamiento de ruido blanco a 20 Hz en lectores disléxicos en el hemisferio izquierdo. En cuanto al procesamiento del habla, Lehongre y sus colaboradores (2013) encontraron que los lectores con dislexia mostraban una menor respuesta oscilatoria durante la visualización de una película con audio en el hemisferio izquierdo.
En general, los estudios publicados han investigado la sincronización entre los ritmos del habla y las oscilaciones neuronales en múltiples bandas de frecuencia (delta, theta y beta / gamma), utilizando diversos estímulos (lingüísticos y no lingüísticos), técnicas de registro (EEG, MEG, fMRI / EEG, fNIRS) y diferentes medidas (PAEee, concentración HbO/HbR, correlación entre la señal BOLD y EEG). Por lo tanto, es muy difícil tener una visión clara de hasta qué punto los resultados publicados en esta área son consistentes, inconsistentes o simplemente no se pueden comparar.
En general, el objetivo de nuestro estudio es comprender mejor las bases oscilatorias neurales subyacentes a las dificultades fonológicas en la dislexia. En primer lugar, identificaremos la (s) banda (s) de frecuencia específica (s) interrumpida (s) en la dislexia. En segundo lugar, aclararemos si las dificultades de sincronización en la dislexia están presentes tanto para estímulos lingüísticos (el habla) como para estímulos no lingüísticos (ruido blanco).
Asimismo, si bien muchos estudios han investigado los mecanismos neuronales que subyacen la segmentación del habla en adultos con dislexia, dichos mecanismos necesitan ser evaluados con mayor profundidad en niños con dislexia. En nuestros estudios, evaluaremos por primera vez tanto a niños como a adultos en tareas auditivas. Esto nos permitirá estudiar el desarrollo de los mecanismos neuronales de segmentación en lectores con y sin dislexia.
Mecanismos neuronales de segmentación del habla en niños con dislexia
Por media de la magnetoencefalografía (MEG), registramos la actividad cerebral de niños y adultos con y sin dislexia mientras escuchaban el habla y una serie de estímulos no lingüísticos (ruido blanco modulado en amplitud a 2, 5 y 30 Hz) (Lizarazu et al., 2015; Molinaro et al., 2016).
Durante el procesamiento del habla, observamos que los lectores disléxicos (tanto niños como adultos) presentan dificultades en la sincronización entre los ritmos del habla y las oscilaciones neuronales en la banda delta (0.5 21 Hz) (Molinaro et al., 2016). La falta de sincronización se localiza en regiones auditiva del hemisferio derecho y en regiones frontales (giro frontal inferior) del hemisferio izquierdo. Estos resultados indican que los problemas de sincronización a bajas frecuencias ocurren en regiones auditivas, pero también en regiones cerebrales asociadas a procesamientos cognitivos de mayor nivel. Además, utilizando el análisis de conectividad causal, demostramos que los problemas de sincronización en las regiones auditivas podrían afectar regiones de mayor jerarquía cognitiva involucradas en el procesamiento del habla.
Los participantes también escuchaban ruido blanco modulado a 2, 4, 7, 30 y 60 Hz. Estas frecuencias de modulación corresponden a unidades fonológicas relevantes en el habla (2 Hz, prosodia; 4 Hz, palabras; 7 Hz, sílabas; 30 y 60 Hz, fonemas). Nuestros resultados indican que los disléxicos (tanto niños como adultos) muestran una sincronización cerebral atípica durante el procesamiento de las modulaciones a 4 Hz y 30 Hz (Lizarazu et al., 2015). También encontramos diferencias entre lectores controles y disléxicos en los valores de lateralización hemisférica durante el procesamiento del ruido blanco modulado a 4 y a 30 Hz. En línea con la teoría de Poeppel (2003), observamos que los controles muestran una lateralización en el hemisferio derecho a los 4 Hz. Sin embargo, los disléxicos no presentan ninguna lateralización a los 4 Hz. A los 30 Hz, observamos un procesamiento bilateral en los controles, mientras que los disléxicos exhiben una lateralización hacia el hemisferio derecho.
Nuestros resultados están en línea con los estudios que muestran que los lectores disléxicos presentan problemas de sincronización entre los ritmos del habla y las oscilaciones neuronales. Observamos que la sincronización neuronal en las bandas delta, theta y gamma son atípicas en los niños y adultos con dislexia. Además, encontramos que los problemas de sincronización ocurren para los estímulos lingüísticos (habla), pero también para los no lingüísticos (ruido blanco). Esto indica que los problemas de sincronización podrían ocurrir a un nivel perceptual auditivo.
Esta y otras evidencias empíricas sugieren que el modelo de sincronización de las ondas cerebrales es prometedor para explicar la calidad en la segmentación e identificación de fonemas. Queda aún camino por recorrer en cuanto a la detección y prevención de las dificultades en el desarrollo de la lectura, pero los estudios actuales son prometedores en cuanto a proveer herramientas para que cada vez menos niños sufran las consecuencias de una lectura ineficaz.
BIBLIOGRAFÍA
- Abrams, D. A., Nicol, T., Zecker, S., & Kraus, N. (2009). Abnormal cortical processing of the syllable rate of speech in poor readers. Journal of Neuroscience, 29(24), 7686-7693.
- Amitay, S., Ahissar, M., & Nelken, I. (2002). Auditory processing deficits in reading disabled adults. Journal of the Association for Research in Otolaryngology, 3(3), 302-320.
- Arvaniti, A. (2009). Rhythm, timing and the timing of rhythm. Phonetica, 66(1-2), 46-63.
- Bourguignon, M., De Tiege, X., de Beeck, M. O., Ligot, N., Paquier, P., Van Bogaert, P., … & Jousmäki, V. (2013). The pace of prosodic phrasing couples the listener’s cortex to the reader’s voice. Human brain mapping, 34(2), 314-326.
- Cutini, S., Szűcs, D., Mead, N., Huss, M., & Goswami, U. (2016). Atypical right hemisphere response to slow temporal modulations in children with developmental dyslexia. Neuroimage, 143, 40-49.
- De Martino, S., Espesser, R., Rey, V., & Habib, M. (2001). The “temporal processing deficit” hypothesis in dyslexia: New experimental evidence. Brain and cognition, 46(1-2), 104-108.
- De Vos, A., Vanvooren, S., Vanderauwera, J., Ghesquière, P., & Wouters, J. (2017). Atypical neural synchronization to speech envelope modulations in dyslexia. Brain and language, 164, 106-117.
- Finucci, J. M., & Childs, B. (1981). Are there really more dyslexic boys than girls. Sex differences in dyslexia, 1-9.
- Geiser, E., Zaehle, T., Jancke, L., & Meyer, M. (2008). The neural correlate of speech rhythm as evidenced by metrical speech processing. Journal of Cognitive Neuroscience, 20(3), 541-552.
- Ghitza, O., & Greenberg, S. (2009). On the possible role of brain rhythms in speech perception: intelligibility of time-compressed speech with periodic and aperiodic insertions of silence. Phonetica, 66(1-2), 113-126.
- Giraud, A. L., & Poeppel, D. (2012). Cortical oscillations and speech processing: emerging computational principles and operations. Nature neuroscience, 15(4), 511.
- Goswami, U. (2011). A temporal sampling framework for developmental dyslexia. Trends in cognitive sciences, 15(1), 3-10.
- Goswami, U., Thomson, J., Richardson, U., Stainthorp, R., Hughes, D., Rosen, S., & Scott, S. K. (2002). Amplitude envelope onsets and developmental dyslexia: A new hypothesis. Proceedings of the National Academy of Sciences, 99(16), 10911-10916.
- Greenberg, S., & Arai, T. (2001). The relation between speech intelligibility and the complex modulation spectrum. In Seventh European Conference on Speech Communication and Technology.
- Gross, J., Hoogenboom, N., Thut, G., Schyns, P., Panzeri, S., Belin, P., & Garrod, S. (2013). Speech rhythms and multiplexed oscillatory sensory coding in the human brain. PLoS biology, 11(12), e1001752.
- Hämäläinen, J. A., Rupp, A., Soltész, F., Szücs, D., & Goswami, U. (2012). Reduced phase locking to slow amplitude modulation in adults with dyslexia: an MEG study. Neuroimage, 59(3), 2952-2961.
- Hickok, G., & Poeppel, D. (2007). The cortical organization of speech processing. Nature reviews neuroscience, 8(5), 393.
- Lehongre, K., Ramus, F., Villiermet, N., Schwartz, D., & Giraud, A. L. (2011). Altered low-gamma sampling in auditory cortex accounts for the three main facets of dyslexia. Neuron, 72(6), 1080-1090.
- Lehongre, K., Morillon, B., Giraud, A. L., & Ramus, F. (2013). Impaired auditory sampling in dyslexia: further evidence from combined fMRI and EEG. Frontiers in human neuroscience, 7, 454.
- Lizarazu, M., Lallier, M., Molinaro, N., Bourguignon, M., Paz‐Alonso, P. M., Lerma‐Usabiaga, G., & Carreiras, M. (2015). Developmental evaluation of atypical auditory sampling in dyslexia: Functional and structural evidence. Human brain mapping, 36(12), 4986-5002.
- Molinaro, N., Lizarazu, M., Lallier, M., Bourguignon, M., & Carreiras, M. (2016). Out‐of‐synchrony speech entrainment in developmental dyslexia. Human brain mapping, 37(8), 2767-2783.
- Molinaro, N., & Lizarazu, M. (2018). Delta (but not theta)‐band cortical entrainment involves speech‐specific processing. European Journal of Neuroscience, 48(7), 2642-2650.
- Panzeri, S., Brunel, N., Logothetis, N. K., & Kayser, C. (2010). Sensory neural codes using multiplexed temporal scales. Trends in neurosciences, 33(3), 111-120.
- Poelmans, H., Luts, H., Vandermosten, M., Boets, B., Ghesquière, P., & Wouters, J. (2012). Auditory steady state cortical responses indicate deviant phonemic-rate processing in adults with dyslexia. Ear and hearing, 33(1), 134-143.
- Poeppel, D. (2003). The analysis of speech in different temporal integration windows: cerebral lateralization as ‘asymmetric sampling in time’. Speech communication, 41(1), 245-255.
- Power, A. J., Mead, N., Barnes, L., & Goswami, U. (2013). Neural entrainment to rhythmic speech in children with developmental dyslexia. Frontiers in human neuroscience, 7, 777.
- Power, A. J., Colling, L. J., Mead, N., Barnes, L., & Goswami, U. (2016). Neural encoding of the speech envelope by children with developmental dyslexia. Brain and language, 160, 1-10.
- Ramus, F., Rosen, S., Dakin, S. C., Day, B. L., Castellote, J. M., White, S., & Frith, U. (2003). Theories of developmental dyslexia: insights from a multiple case study of dyslexic adults. Brain, 126(4), 841-865.
- Ramus, F. (2004). Neurobiology of dyslexia: A reinterpretation of the data. TRENDS in Neurosciences, 27(12), 720-726.
- Rey, V., De Martino, S., Espesser, R., & Habib, M. (2002). Temporal processing and phonological impairment in dyslexia: Effect of phoneme lengthening on order judgment of two consonants. Brain and language, 80(3), 576-591.
- Saberi, K., & Perrott, D. R. (1999). Cognitive restoration of reversed speech. Nature, 398(6730), 760.
- Tallal, P. (1980). Language and reading: Some perceptual prerequisites. Bulletin of the Orton Society, 30(1), 170-178.
- Vellutino, F. R., Fletcher, J. M., Snowling, M. J., & Scanlon, D. M. (2004). Specific reading disability (dyslexia): What have we learned in the past four decades?. Journal of child psychology and psychiatry, 45(1), 2-40.
- Wagner, R. K., & Torgesen, J. K. (1987). The nature of phonological processing and its causal role in the acquisition of reading skills. Psychological bulletin, 101(2), 192.
DECLARACIÓN DE TRANSPARENCIA
Los autores declaran no tener algún conflicto de intereses, o potencial conflicto de intereses, que pueda sesgar su trabajo, o pudiera ser percibido como un sesgo en su trabajo, así como agradecer todo el apoyo financiero y colaboraciones personales.
Todos los pacientes incluidos en el estudio dieron su consentimiento a participar después de haber sido informados de forma concienzuda acerca del estudio.
Nicola Molinaro
Real Academia Nacional de Medicina de España
C/ Arrieta, 12 · 28013 Madrid
Tlf.: +34 91 159 47 34 | Email de correspondencia
Año 2018 · 135 (02) · Supl. 01 · páginas 47 a 51
Enviado*: 23.05.18
Revisado: 30.05.18
Aceptado: 25.06.18
* Fecha de lectura en la RANM