MECANISMOS DE PLASTICIDAD SINÁPTICA Y NEUROMODULACIÓN: IMPLICACIONES EN ENFERMEDADES NEUROLÓGICAS
MECHANISMS OF SYNAPTIC PLASTICITY AND NEUROMODULATION: IMPLICATION IN NEUROLOGICAL DISEASES
Sandra Jurado Sánchez
Instituto de Neurociencias CSIC-UMH, Alicante
Los trastornos sociales son un síntoma comórbido común de varias afecciones patológicas, como el síndrome del espectro autista y enfermedades neurodegenerativas como la enfermedad de Alzheimer. Los trastornos sociales se caracterizan por su heterogeneidad sintomática que incluye déficits en la interacción y comunicación social, conductas repetitivas y alteraciones cognitivas que tienen un alto impacto en la calidad de vida. Debido a su amplio espectro, los déficits sociales se consideran una preocupación médica creciente, ya que el número de personas afectadas ha aumentado rápidamente en los últimos años, exponiendo un preocupante escenario. Además, la agresividad y las respuestas sociales aberrantes son síntomas comunes de enfermedades neurodegenerativas como la enfermedad de Alzheimer, de creciente incidencia en sociedades cada vez más envejecidas. Sin embargo, los mecanismos por los cuales los procesos neurodegenerativos afectan a la regulación de la interacción social han sido poco estudiados.
Actualmente no existen tratamientos efectivos para los trastornos de la conducta social, aunque la oxitocina (OXT), un neuropéptido endógeno involucrado en el estrés y la interacción social, se ha identificado como una potencial diana terapéutica para aliviar déficits sociales. Ensayos clínicos basados en la administración exógena de OXT han producido resultados prometedores pero inconsistentes, probablemente debido a la baja permeabilidad de la OXT administrada exógenamente (por ejemplo, intranasal) a través de la barrera hematoencefálica. Estos resultados indican la necesidad de explorar estrategias alternativas para estimular el sistema oxitocinérgico endógeno, ya sea activando el receptor de OXT o induciendo su liberación en el sistema nervioso central (Figura 1). A pesar de su importancia para regular funciones homeostáticas y comportamientos complejos, aún se desconocen muchos detalles sobre los mecanismos que regulan la liberación de OXT en el cerebro. Esta falta de información contrasta con el amplio conocimiento disponible sobre los mecanismos implicados en la exocitosis de neurotransmisores clásicos como el glutamato o GABA. En gran medida, este escenario ha sido provocado por la falta de herramientas moleculares que permitan la identificación y estudio de vesículas de núcleo denso, que almacenan diferentes neuromoduladores como la OXT. Nuestro laboratorio ha implementado una estrategia multidisciplinar basada en nuevas técnicas de clarificación de tejido iDISCO+, imagen de ultra resolución 3D, electrofisiología y optogenética para analizar las propiedades y mecanismos reguladores del sistema oxitocinérgico durante el desarrollo y el envejecimiento. Nuestro trabajo ha permitido identificar moléculas clave implicadas en la secreción de OXT. Este conocimiento constituye la base para desarrollar nuevas herramientas para regular la liberación de neuromoduladores como la OXT en el sistema nervioso central. Además, estas nuevas herramientas permitirán analizar el papel de la OXT en la plasticidad sináptica, una propiedad neuronal que permite la adaptación de los circuitos neuronales, y que se encuentra alterada durante trastornos neurodegenerativos.

Social disorders are a common comorbid symptom of several pathological conditions, such as autism spectrum syndrome and neurodegenerative diseases such as Alzheimer’s disease. Social disorders are characterized by their symptomatic heterogeneity including deficits in social interaction and communication, repetitive behaviors and cognitive alterations that have a high impact on quality of life. Given their broad spectrum, social deficits are considered a growing medical concern since the number of people affected by social disorders such as social anxiety has increased rapidly in recent years exposing a concerning scenario. Furthermore, aggressiveness and aberrant social responses are common symptoms of neurodegenerative disorders such as Alzheimer’s disease, of growing incidence in increasingly aging societies. However, the mechanisms by which neurodegeneration affects the regulation of social interaction have been scarcely studied.
Currently there are no effective treatments for social behavior disorders, however oxytocin (OXT), an endogenous neuropeptide involved in stress and social interaction, it has recently been identified as a potential therapeutic target to alleviate social deficits. Clinical trials based on OXT´s exogenous administration have yielded promising but inconsistent results, probably due to the low permeability of exogenously administered (e.g. intranasal) OXT across the blood-brain barrier. These results indicate the need to explore alternative strategies to stimulate the endogenous oxytocinergic system either by activating the OXT receptor or inducing its release in the central nervous system (Figure 1). Despite its importance in regulating homeostatic functions and complex behaviors, many details about the mechanisms that regulate OXT release in the brain are still unknown. This lack of information contrasts with the extensive knowledge available on the mechanisms involved in the exocytosis of classical neurotransmitters such as glutamate or GABA. This scenario is partly due to the lack of molecular tools to unambiguously identify and study dense core vesicles, which store many different brain neuromodulators such as OXT. Our laboratory has implemented a multidisciplinary strategy based on novel iDISCO+ tissue clarification techniques, 3D ultra-resolution imaging, electrophysiology, and optogenetics to analyze the properties and regulatory mechanisms of the oxytocinergic system during development, natural and pathological aging. Our work has revealed key molecules involved in OXT secretion, which has allowed us to develop new tools to regulate the release of neuromodulators such as OXT in the central nervous system to analyze their role in synaptic plasticity, a neuronal property that allows neuronal circuit adaptation, and which is altered during neurodegeneration.

DESARROLLO Y PLASTICIDAD DE LOS SISTEMAS DE OXITOCINA Y VASOPRESINA EN EL HIPOTÁLAMO
DEVELOPMENT AND PLASTICITY OF THE HYPOTHALAMIC OXYTOCINERGIC AND VASOPRESSINERGIC SYSTEMS
M.P. Madrigal, S. Jurado
Instituto de Neurociencias CSIC-UMH, Alicante
Los neuropéptidos oxitocina (OXT) y arginina vasopresina (AVP) juegan un papel crítico en la regulación de funciones homeostáticas y comportamientos complejos como la interacción social. Alteraciones en la función de estos sistemas se han relacionado con déficits cognitivos y sociales, sin embargo, aún se desconocen muchos detalles sobre el desarrollo y la especificación de estos circuitos neuronales. Ambos neuropéptidos se sintetizan principalmente en núcleos hipotalámicos como el núcleo paraventricular y supraóptico (PVN y SON).
Nuestro estudio ha implementado novedosas técnicas de clarificación de tejido (iDISCO+) y microscopía de ultraresolución 3D (Figura 1A) para analizar la formación de estos circuitos durante el desarrollo del cerebro del ratón y las propiedades de plasticidad celular en respuesta a periodos críticos como la maternidad o la experiencia sexual en el animal adulto. Nuestros resultados indican que durante la etapa postnatal temprana existe un elevado número de neuronas hipotalámicas que coexpresan OXT y AVP, coincidiendo con un período crítico para la interacción social. Esta población mixta disminuye drásticamente en el cerebro adulto, lo que sugiere que la maduración del sistema requiere de la activación de procesos de plasticidad celular (Figura 1B). Reconstrucciones 3D del cerebro adulto han revelado que las neuronas de OXT y AVP en el área retroquiasmática (RCH) y el núcleo accesorio (AN) también experimentan fenómenos de plasticidad celular en respuesta a la experiencia sexual y la maternidad. Nuestras observaciones indican una prevalencia de neuronas AVP en el SON de hembras vírgenes que se convierten en neuronas positivas a OXT después de dar a luz, al contrario de lo observado en el AN.
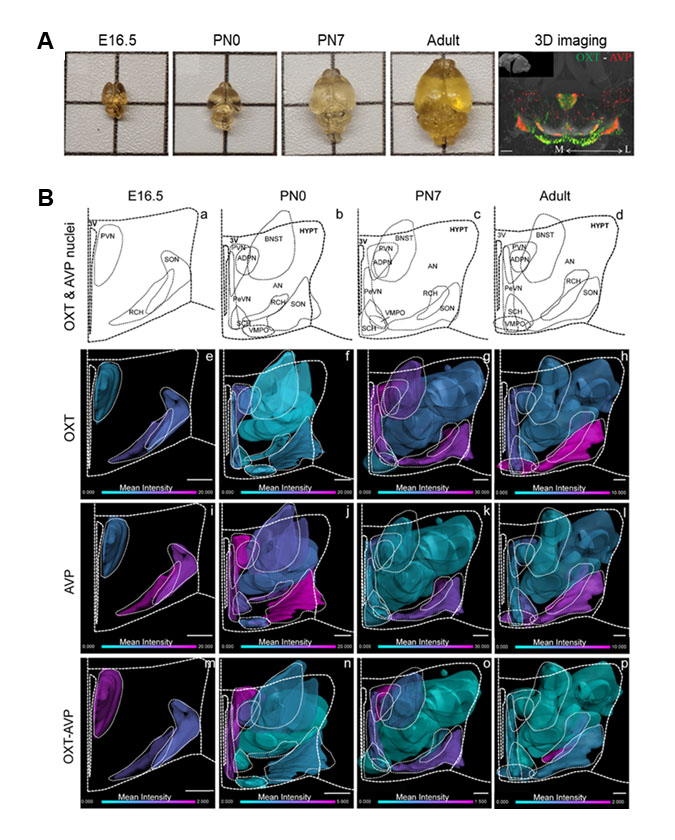
Por otra parte, hemos analizado otros tipos neuronales en el hipotálamo como neuronas inhibitorias positivas a GAD67 y neuronas dopaminérgicas identificadas mediante el marcador de monoamina tirosina hidroxilasa (TH). Nuestros resultados indican la presencia de neuronas positivas para GAD67 en el hipotálamo, pero una mínima expresión en células de OXT o AVP. Además, encontramos que la experiencia sexual induce la expresión de TH en una subpoblación de neuronas de AVP y OXT en el RCH. Nuestros hallazgos proporcionan nueva información básica para comprender la especificación de los sistemas de OXT y AVP durante el desarrollo y sus propiedades plásticas durante periodos críticos en el animal adulto.
The neuropeptides oxytocin (OXT) and arginine vasopressin (AVP) play critical roles in the regulation of homeostatic functions and complex behaviors such as social interaction. Alterations of these systems have been related to cognitive and social deficits, however, many details about the development and specification of these neural circuits remain unknown. Both neuropeptides are mainly synthesized in hypothalamic nuclei such as the paraventricular and supraoptic nucleus (PVN and SON).
Our study has implemented novel tissue clarification techniques (iDISCO+) and 3D ultra-resolution microscopy (Figure 1A) to analyze the formation of these circuits during mouse brain development and the properties of cellular plasticity in response to critical periods such as maternity or sexual experience in the adult animal. Our results indicate that during early postnatal stages there is a high number of hypothalamic neurons that co-express OXT and AVP, coinciding with a critical period for social interaction. This mixed population decreases dramatically in the adult brain, suggesting that the maturation of these systems requires a switch in their internal program which modifies neuropeptide expression via cellular plasticity mechanisms (Figure 1B). Furthermore, 3D reconstruction of the adult brain indicates that OXT and AVP neurons in the retrochiasmatic area (RCH) and the accessory nucleus (AN) also undergo cellular plasticity processes in response to sexual experience and motherhood. Our observations indicate a prevalence of AVP neurons in the SON of naïve females that become OXT-positive neurons after giving birth, contrary to that observed in the AN.
On the other hand, we have analyzed other neuronal types in the hypothalamus such as GAD67-positive inhibitory neurons and dopaminergic neurons identified by the monoamine tyrosine hydroxylase (TH) marker. Our results indicate the presence of GAD67-positive neurons in the hypothalamus, but minimal co-localization in OXT or AVP cells. Furthermore, we found that sexual experience induces TH expression in a subpopulation of AVP and OXT neurons in the RCH. Our findings provide new fundamental information for understanding the specification of OXT and AVP systems during development and their plastic properties in response to critical periods in the adult animal.
DINÁMICA Y MECANISMO MOLECULAR DE LA LIBERACIÓN DE OXITOCINA EN NEURONAS HIPOTALÁMICAS
DYNAMICS AND MOLECULAR MECHANISM OF OXYTOCIN RELEASE IN HYPOTHALAMIC NEURONS
Aznar-Escolano B., Madrigal M.P., Jurado S.
Instituto de Neurociencias CSIC-UMH, Alicante
El sistema oxitocinérgico regula funciones homeostáticas vitales y distintos aspectos de la interacción social como la memoria social. La desregulación de este sistema se ha implicado en desórdenes neurológicos caracterizados por déficits sociales como el trastorno del espectro autista. A pesar de su importante acción neuromoduladora, aún se desconocen muchos detalles sobre las propiedades y el mecanismo de liberación de la oxitocina (OXT) en el Sistema Nervioso Central (SNC).
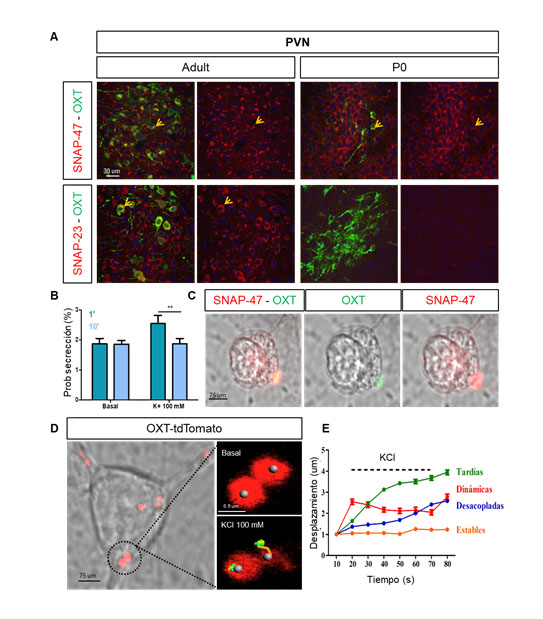
Las neuronas oxitocinérgicas del hipotálamo son las responsables de modular los niveles de OXT en el cerebro principalmente a través de eventos de liberación somatodendrítica. Sin embargo, la maquinaria de fusión responsable de la secreción de OXT no ha sido caracterizada en detalle, principalmente por la carencia de marcadores específicos. El análisis de la expresión de proteínas SNARE y sinaptotagminas, reveló que dos proteínas SNARE: SNAP-47 y SNAP-23 se localizan en el compartimento somatodendrítico de las neuronas oxitocinérgicas, sugiriendo un papel en la exocitosis de OXT (Figura 1A). Nuestros datos indican que la expresión de estas isoformas se encuentra regulada durante el desarrollo. Así, mientras que SNAP-47 está presente en el hipotálamo desde el estadío prenatal, la expresión de SNAP-23 no es detectable hasta la edad adulta.
Técnicas de chasing vesicular en célula única han demuestrado un aumento significativo de la probabilidad de liberación de OXT en respuesta a la estimulación neuronal durante los primeros segundos (60 s), mientras que tiempos de estimulación prolongados (p.ej. 10 min) no afectan a la probabilidad de liberación sugiriendo una depleción de vesículas de OXT listas para ser liberadas (Figura 1B). Nuestros resultados indican que la exocitosis de OXT preferentemente ocurre en regiones enriquecidas con SNAP-47 (Figura 1C), a través un mecanismo parcialmente independiente de la entrada de calcio. Paralelamente se ha realizado un análisis exhaustivo de la dinámica y propiedades de liberación de OXT en neuronas hipotalámicas en cultivo combinando novedosas técnicas de imagen con la línea del ratón transgénico OXT-tdTomato (Figura 1D), que permite la identificación de las vesículas de OXT. La visualización de vesículas somatodendríticas de OXT ha revelado la existencia de poblaciones vesiculares con diferentes dinámicas en respuesta a la estimulación neuronal. Se identificaron cuatro grupos principales: i) vesículas dinámicas, las cuales se movilizan durante los primeros 10 s de estimulación, ii) vesículas tardías, el grupo más abundante, con cambios de desplazamiento tras 20-40 s de estimulación (Figura 1E); iii) vesículas desacopladas, las cuales experimentan cambios de movilidad durante el periodo final de estimulación; y iv) vesículas estables (sin movilidad detectable). Registros electrofisiológicos preliminares indican que las neuronas oxitocinérgicas en cultivo presentan cinéticas lentas de despolarización, lo que podría subyacer la dinámica tardía de las vesículas de OXT y su dependencia parcial a la entrada de calcio. Nuestros resultados han revelado la existencia de diferentes grupos de vesículas de OXT, que se movilizan de forma diferente en respuesta al mismo estímulo, una característica hasta ahora desconocida y que puede ser relevante para la modulación fina del comportamiento.
The oxytocinergic system regulates vital homeostatic functions and different aspects of social interaction such as social bonding or social memory. Dysregulation of this system has been implicated in neurological disorders characterized by social deficits such as autism spectrum disorder. Despite its important neuromodulatory action, many details about the properties and release mechanism of oxytocin (OXT) in the Central Nervous System (CNS) are still unknown.
Oxytocinenergic neurons in the hypothalamus are responsible for modulating OXT levels in the brain mainly through somatodendritic release events. However, the fusion machinery responsible for OXT secretion has not been characterized in detail, mainly due to the lack of specific markers for these vesicles in neurons. Analysis of the expression of SNARE and synaptotagmin proteins revealed that two SNARE proteins: SNAP-47 and SNAP-23 are found in the somatodendritic compartment of oxytocinenergic neurons, suggesting a role in OXT exocytosis (Figure 1A). Our data indicate that the expression of these isoforms is developmentally regulated. Thus, whereas SNAP-47 is present in the hypothalamus from the prenatal stage, the expression of SNAP-23 is not detectable until adulthood.
Single cell vesicular chasing techniques have demonstrated a significant increase in the probability of OXT release in response to neuronal stimulation during the first seconds of stimulation (60 s), whereas longer stimulation times (10 min) do not affect the release probability suggesting a depletion of OXT vesicles ready to be released (Figure 1B). Our results indicate that OXT exocytosis occurs preferentially in regions enriched with SNAP-47 (Figure 1C), through a mechanism partially independent of calcium influx. In parallel, an exhaustive analysis of the dynamic properties of OXT release in cultured hypothalamic neurons has been carried out combining novel imaging techniques and the transgenic mouse line OXT-tdTomato (Figure 1D), which allows the unambiguous identification of OXT vesicles. Visualization of somatodendritic OXT vesicles has revealed the existence of distinct vesicular populations with different dynamics in response to neuronal stimulation. Four main groups have been identified: i) dynamic vesicles, mobilized during the first 10 s of stimulation, ii) delayed vesicles, the most abundant group, with displacement changes after 20-40 s of stimulation (Figure 1E); iii) uncoupled vesicles, which undergo mobility changes during the final period of stimulation; and iv) stable vesicles (no detectable mobility). Preliminary electrophysiological recordings indicate that cultured oxytocinergic neurons show slow depolarization kinetics, which could underlie the slow dynamics of OXT vesicles and their partial dependence to calcium influx. Our results revealed the existence of different groups of OXT vesicles, which are distinctly mobilized in response to the same stimulus, exposing an unknown feature that may be relevant for the fine tuning of behavior.